6.3. Роль различных отделов цнс в регуляции движений
Спинной мозг обеспечивает протекание многих элементарных двигательных рефлексов, включение которых в сложные двигательные акты и регуляция по мощности, пространственной ориентации и моменту включения осуществляется вышележащими отделами головного мозга под контролем коры больших полушарий.
6.3.1. Роль спинного мозга и подкорковых отделов ЦНС в регуляции движений
Спинной мозг осуществляет ряд элементарных двигательных рефлексов: рефлексы на растяжение (миотатические и сухожильные рефлексы, например коленный рефлекс), кожные сгибательные рефлексы(например защитный рефлекс отдергивания конечности при уколах, ожогах), разгибательные рефлексы(рефлекс отталкивания от опоры, лежащий в основе стояния, ходьбы, бега), перекрестные рефлексы и др.
Элементарные двигательные рефлексы включаются в более сложные двигательные акты – регуляцию деятельности мышц-антагонистов, ритмических и шагательных рефлексов, лежащих в основе локомоций и других движений.
Для сгибательного движения в суставе необходимо не только сокращение мышц-сгибателей, но и одновременное расслабление мышц-разгибателей. При этом в мотонейронах мышц-сгибателей возникает процесс возбуждения, а в мотонейронах мышц-разгибателей – торможение. При разгибании сустава, наоборот, тормозятся центры сгибателей и возбуждаются центры разгибателей. Такие координационные взаимоотношения между спинальными моторными центрами названы реципрокной (взаимосочетанной) иннервацией мышц-антагонистов. Однако реципрокные отношения между центрами мышц-антагонистов в необходимых ситуациях (например, при фиксации суставов, при точностных движениях) могут сменяться одновременным их возбуждением.
Составной частью различных сложных двигательных действий, как произвольных, так и непроизвольных, часто являются ритмические рефлексы. Это одна из форм древних и относительно простых рефлексов. Они особенно выражены при выполнении циклической работы, включаются в шагательные рефлексы. Основные механизмы шагательных движений заложены в спинном мозге. Специальные нейроны (спинальные локомоторные генераторы)и многочисленные взаимосвязи внутри спинного мозга обеспечивают последовательную активность различных мышц конечностей, согласование ритма и фаз движений, приспособление движений к нагрузке на мышцы. В среднем мозгу расположены нейроны "локомоторной области",которые включают этот механизм и регулируют мощность работы мышц, обеспечивая примитивную форму локомоции – без ориентации в пространстве.
Нейроны промежуточной продольной зоны коры мозжечка согласуют лозные реакции с движениями. Они выполняют также точные расчеты по ходу движений, необходимые для коррекции ошибок и адаптации моторных программ к текущей ситуации.Программирование каждого последующего шага осуществляется ими на основе анализа предыдущего. Кроме того, производится согласование движений рук и ног, и особенно – регуляция активности мышц-разгибателей, обеспечивающих опорную фазу движения. Значение мозжечка в четком поддержании темпа ритмических движенийобъясняют геометрически правильным чередованием рядов эфферентных клеток Пуркинье и походящих к ним афферентных волокон.
К управлению ритмическими движениями непосредственное отношение имеют активирующие и угнетающие отделы ретикулярной формации,влияющие на силу и темп сокращения мышц, а также подкорковые ядра,которые организуют автоматическое их протекание и содружественные движения конечностей. Включение древних форм ритмических движений (циклоидных)в акт письма позволяет человеку перейти от отдельного начертания букв к обычной письменной скорописи. То же самое происходит при освоении акта ходьбы – с переходом от отдельных шагов к ритмической походке. Плавность ритмических движений,четкое чередование реципрокных сокращений мышц обеспечивают премоторные отделы коры.
6.3.2. Роль различных отделов коры больших полушарий
Функцией комплекса различных корковых областейявляется определение целесообразности локомоций, их смысла, ориентации в пространстве, перестройка программ движений в различных ситуациях, включение ритмических движений как составного элемента в сложные акты поведения. Об участии различных корковых областей в регуляции циклических движений можно судить по появлению в их электрической активности медленных потенциалов в темпе движения – "меченых ритмов" ЭЭГ, а при редких движениях – по изменениям огибающей амплитуду ЭЭГ кривой.
В организации двигательных актов участвуют практически все отделы коры больших полушарий.Моторная область коры (прецентральная извилина) посылает импульсы к отдельным мышцам,преимущественно к дистальным мышцам конечностей. Объединение отдельных элементов движения в целостный акт ("кинетическую мелодию") осуществляют вторичные поля премоторной области. Они определяют последовательность двигательных актов,формируют ритмические серии движений, регулируют тонус мышц. Постцентральная извилина коры представляет собой общечувствительное поле, которое обеспечивает субъективное ощущение движений.Нижнетеменные области коры (задние третичные поля) формируют представления о взаимном расположении различных частей тела и положении тела в пространстве, обеспечивают точную адресацию моторных команд к отдельным мышцам и пространственную ориентацию движений. Области коры, относящиеся к лимбической системе (нижние и внутренние части коры), ответственны за эмоциональную окраску движенийи управление вегетативными их компонентами.
В высшей регуляции произвольных движений важнейшая роль принадлежитпереднелобным областям (передним третичным полям). Здесь помимо обычных вертикальных колонок нейронов существует принципиально новый тип функциональной единицы – в форме замкнутого нейронного кольца. Циркуляция импульсов в этой замкнутой системе обеспечивает кратковременную память. Она сохраняет в коре возбуждение между временем прихода сенсорных сигналов и формированием ответной эфферентной команды. Такой механизм служит основой сенсомоторной интеграции при программировании движений, при осуществлении зрительно-двигательных реакций.
Функцией переднелобной (третичной) области корыявляется сознательная оценка текущей ситуации и предвидение возможного будущего, выработка цели и задачи поведения, программирование произвольных движений, их контроль и коррекция. Соответствие выполняемых действий поставленным задачам придает движениям человека определенную целесообразность и осмысленность. При поражении лобных долей движения человека становятся бессмысленными.
6.3.3. Речевая регуляция движений
Спецификой регуляции движений у человека является то, что они подчиненыречевым воздействиям, т. е. могут программироваться лобными долями в ответ на поступающие извне словесные сигналы, а также благодаря участию внешней или внутренней речи (мышления) самого человека. В этой функции принимают участие расположенные в левом полушарии человека сенсорный центр речи Вернике и моторный центр речи – центр Брока. Считают, что афферентная импульсация от речевой мускулатуры является важным ориентиром, дополняющим проприоцептивные сигналы от работающих мышц, а формирующиеся на речевой основе избирательные связи в коре облегчают составление моторных программ.
Эта управляющая система еще не развита у ребенка 2–3 лет. Она появляется лишь к 3–4 годам. Внешняя речь, сменяясь постепенно шепотом и переходя затем во внутреннюю речь, становится важным регулятором моторных действий взрослого человека.
6.4. Нисходящие моторные системы
Высшие отделы головного мозга осуществляют свои влияния на деятельность нижележащих отделов, в том числе спинного мозга, через нисходящие пути, которые группируют обычно в две основные нисходящие системы – пирамидную и экстрапирамидную.
В настоящее время предлагают подразделять основные нисходящие пути, исходя из расположения нервных окончаний в спинном мозге и функциональных различий, на следующие две системы: более молодую латеральную, волокна которой оканчиваются в боковых (латеральных) частях спинного мозга и связанную преимущественно с мускулатурой дистальных звеньев конечностей(сюда относят корково-спинномозговую и красно-ядерно-спинномозговую системы), и древнюю медиальную, волокна которой оканчиваются во внутренних (медиальных) частях белого вещества, связанную главным образом с мускулатурой туловища и проксимальных звеньев конечностей,состоящую из вестибуло-спинномозговой и ретикуло-спинномозговой систем.
Пирамидная система выполняет три основные функции:
• посылает мотонейронам спинного мозга импульсы – команды к движениям (пусковые влияния);
• изменяет проведение нервных импульсов во вставочных спинальных нейронах,облегчая протекание нужных в данный момент спинномозговых рефлексов;
• осуществляет контроль за потоками афферентных сигналовв нервные центры, выключая постороннюю информацию и обеспечивая обратные связи от работающих мышц.
Волокна пирамидной системы вызывают преимущественно возбуждение мотонейронов мышц-сгибателей, особенно влияя на отдельные мышцы и даже части мышц верхних конечностей, в частности на мышцы пальцев рук.
Экстрапирамидная система оказывает обобщенные воздействия на познотонические реакции организма от коры, мозжечка, промежуточного мозга и подкорковых ядер. Влияния этой системы передаются через корково-красноядерно-спинномозговой путь, составляющий функционально единое целое с "медленной" подсистемой пирамидного тракта, и через более древнюю медиальную систему (вестибуло-спинномозговую и ретикуло-спинномозговую системы).
Таким образом, среди нисходящих моторных систем, осуществляющих функцию контроля активности мотонейронов спинного мозга, можно выделить одну часть, которая обусловливает фазную двигательную деятельность – это "быстрая"· подсистема пирамидного тракта, и другую часть – остальные нисходящие системы, которые обеспечивают регуляцию тонуса мышц и позных реакций организма. Из этих систем три системы обеспечивают повышение возбудимости мотонейронов мышц-сгибателей (корково-спинномозговая, корково-красноядерно-спинномозговая и корково-ретикуло-спинномозговая), а одна система (вестибуло-спиномозговая) тормозит эти мотонейроны (рис. 15).
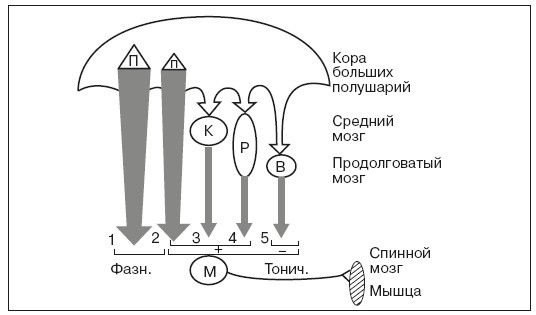
Рис. 15. Схема основных нисходящих путей регуляции двигательной деятельности:
1 – быстрая подсистема и 2 – медленная подсистема корково-спинномозгового пути (пирамидного тракта); 3 – корково-красноядерно-спинномозговой путь. Латеральная система – 1, 2, 3. Медиальная система – 4, 5. М – мотонейрон спинного мозга, получающий фазные (Фазн.) и тонические (Тонич.) возбуждающие (+) и тормозящие (-) влияния; П – пирамидная система: К – красная ядра; Р – ретикулярная система; В – вестибулярные ядра
7. Сенсорные системы
Сложные акты поведения человека во внешней среде требуют постоянного анализа окружающего мира, а также осведомленности нервных центров о состоянии внутренних органов. Специальные нервные аппараты, служащие для анализа внешних и внутренних раздражений, И.П. Павлов назвал анализаторами. Современное представление об анализаторах как сложных многоуровневых системах, передающих информацию от рецепторов к коре и включающих регулирующие влияния коры на рецепторы и нижележащие центры, привело к появлению более общего понятия – сенсорные системы.
7.1. Общий план организации и функции сенсорных систем
В составе сенсорной системы различают три отдела:
1) периферический, состоящий из рецепторов, воспринимающих определенные сигналы, и специальных образований, способствующих работе рецепторов (эта часть представляет собой органы чувств – глаз, ухо и др.); 2) проводниковый, включающий проводящие пути и подкорковые нервные центры;
3) корковый – области коры больших полушарий, которым адресуется данная информация.
Нервный путь, связывающий рецептор с корковыми клетками, обычно состоит из четырех нейронов: первый чувствительный нейронрасположен вне ЦНС – в спинномозговых узлах или узлах черепномозговых нервов (спиральном узле улитки, вестибулярном узле и др.); второй нейроннаходится в спинном, продолговатом или среднем мозге; третий нейрон – в релейных (переключательных) ядрах таламуса (промежуточный мозг); четвертый нейронпредставляет собой корковую клетку проекционной зоны коры больших полушарий.
Основные функции сенсорных систем:
сбор и обработка информациио внешней и внутренней среде организма;
осуществление обратных связей,информирующих нервные центры о результатах деятельности;
поддержание нормального уровня (тонуса) функционального состояния мозга.
Разложение сложностей внешнего и внутреннего мира на отдельные элементы и их анализ И.П. Павлов считал основной функцией сенсорных систем (анализаторов). Помимо первичного сбора информацииважной функцией сенсорных систем является также осуществление обратных связей о результатах деятельности организма. Для уточнения и совершенствования различных действий человека, в первую очередь двигательных, ЦНС должна получать информацию о силе и длительности выполняемых сокращений мышцами, о скорости и точности перемещений тела или рабочих снарядов, об изменениях темпа движений, о степени достижения поставленной цели и т. п. Без этой информации невозможно формирование и совершенствование двигательных навыков, в том числе спортивных, затруднено совершенствование техники выполняемых упражнений.
Наконец, сенсорные системы вносят свой вклад в регуляцию функционального состояния организма.Импульсация, идущая от различных рецепторов в кору больших полушарий как по специфическим, так и по неспецифическим путям, является существенным условием поддержания нормального уровня ее функционального состояния. Искусственное выключение органов чувств в специальных экспериментах на животных приводило к резкому снижению тонуса коры и засыпанию. Такое животное просыпалось лишь во время кормления и при позывах к мочеиспусканию или опорожнению кишечника.
7.2. Классификация и механизмы возбуждения рецепторов
Рецепторами называются специальные образования, трансформирующие (преобразующие) энергию внешнего раздражения в специфическую энергию нервного импульса.
Все рецепторы по характеру воспринимаемой средыделятся на экстерорецепторы, принимающие раздражения из внешней среды (рецепторы органов слуха, зрения, обоняния, вкуса, осязания), интерорецепторы, реагирующие на раздражения из внутренних органов, и проприорецепторы, воспринимающие раздражения из двигательного аппарата (мышц, сухожилий, суставных сумок).
По виду воспринимаемых раздраженийразличают хеморецепторы (рецепторы вкусовой и обонятельной сенсорных систем, хеморецепторы сосудов и внутренних органов); механорецепторы (проприорецепторы двигательной сенсорной системы, барорецепторы сосудов, рецепторы слуховой, вестибулярной, тактильной и болевой сенсорных систем); фоторецепторы (рецепторы зрительной сенсорной системы) и терморецепторы (рецепторы температурной сенсорной системы кожи и внутренних органов).
По характеру связи с раздражителемразличают дистантные рецепторы, реагирующие на сигналы от удаленных источников и обусловливающие предупредительные реакции организма (зрительные и слуховые), и контактные, принимающие непосредственные воздействия (тактильные и др.).
По структурным особенностям различают первичные и вторичные рецепторы. Первичные рецепторы – это окончания чувствительных биполярных клеток, тело которых находится вне ЦНС, один отросток подходит к воспринимающей раздражение поверхности, а другой направляется в ЦНС (например, проприорецепторы, терморецепторы, обонятельные клетки). Вторичные рецепторыпредставлены специализированными рецепторными клетками, которые расположены между чувствительным нейроном и точкой приложения раздражителя (например, фоторецепторы глаза).
В первичных рецепторах энергия внешнего раздражителя непосредственно преобразуется в нервный импульс в одной и той же клетке.В периферическом окончании чувствительных клеток при действии раздражителя возникает повышение проницаемости мембраны и ее деполяризация, возникает местное возбуждение – рецепторный потенциал, который, достигнув пороговой величины, обусловливает появление потенциала действия, распространяемого по нервному волокну к нервным центрам.
Во вторичных рецепторах раздражитель вызывает появление рецепторного потенциала в клетке-рецепторе.Ее возбуждение приводит к выделению медиатора в пресинаптической части контакта клетки-рецептора с волокном чувствительного нейрона. Местное возбуждение этого волокна отражается появлением возбуждающего постсинаптического потенциала или так называемого генераторного потенциала. При достижении порога возбудимости в волокне чувствительного нейрона возникает потенциал действия, несущий информацию в ЦНС. Таким образом, во вторичных рецепторах одна клетка преобразует энергию внешнего раздражителя в рецепторный потенциал, а другая – в генераторный потенциал и потенциал действия.