МЕТЭСТРУС
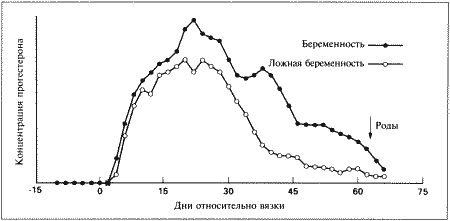
Фаза метэструса следует за эструсом; ее начало определяют по снижению полового возбуждения у самки (обычно на 6–8 день после начала эструса или 8–10 день после преовуляторного пика ЛГ). Переход к метэструсу сопровождается изменением характера вагинальных выделений (повышается содержание лейкоцитов и возрастает количество клеток, содержащих ядра); эндоскопия выявляет сокращение и истончение профиля слизистой влагалища, свидетельствующие об окончании фертильного периода. Специфических изменений эндокринного статуса не наблюдается, за исключением неуклонного, продолжающегося от начала эструса, повышения концентрации прогестерона до 30–90 нмоль/л (10–30 нг/мл) (фиг. 1.2). Как указано выше, беременность развивается в фазе метэструса, т. е. в условиях доминирования прогестерона. Как при наличии, так и в отсутствие беременности прогестерон вырабатывается исключительно желтыми телами; плацента в секреции прогестерона не участвует.
На стадии метэструса происходит развитие эмбрионов; интересно отметить согласованность протекания некоторых основных этапов, предшествующих имплантации, и продемонстрировать их связь с изменениями концентрации прогестерона и эстрадиола (фиг. 1.1). Наиболее целесообразно принять за точку отсчета овуляторный пик ЛГ. Как уже упоминалось, овуляция происходит на 2 день, после чего яйцеклетки проходят период созревания. К 4–7 дню они готовы к оплодотворению. Оплодотворенные яйцеклетки некоторое время удерживаются в маточных трубах, они проходят маточно-трубное соединение и приблизительно на 10 день перемещаются в рога матки. Увеличение соотношения прогестерон/эстрадиол (фиг. 1.1), судя по всему, имеет жизненно важное значение для этого этапа. Бластоцисты локализуются в роге матки вплоть до 13 дня, после чего свободно мигрируют из одного рога в другой до 16 дня, когда набухание эндометрия свидетельствует о том, что имплантация эмбрионов произошла.
С начала метэструса концентрация прогестерона продолжает нарастать, достигая максимума (порядка 90–270 нмоль/л; 30–90 нг/мл) к 30 дню от пика ЛГ, а затем постепенно снижается до 30 нмоль/л (10 нг/мл) к 60 дню от пика ЛГ. Таким образом, концентрация прогестерона в крови сохраняется на уровне 30–90 нмоль/л (10–30 нг/мл) около 2 месяцев после окончания эструса; более низкий уровень указывает на ановуляторный цикл или дисфункцию желтых тел. Вплоть до 60 дня время и диапазон колебаний концентрации прогестерона у беременных и небеременных самок одинаковы. Различия проявляются в резком снижении концентрации прогестерона перед родами (см. ниже), тогда как у небеременных самок, у которых отсутствует лютеолитический механизм, концентрация прогестерона снижается постепенно и достигает 3–9 нмоль/л (1–3 нг/мл) в течение последующих 30–60 дней. Неизвестно, проявляется ли активность желтых тел в строго запрограммированный отрезок времени или спад активности объясняется снижением лютеотрофической поддержки.
Повышение концентрации пролактина в сыворотке, отмечающееся как у беременных, так и у небеременных сук в период с 30 по 65 день после пика ЛГ, сохраняется до 60–90 дня, что особенно характерно для кормящих сук, поскольку пролактин обеспечивает секрецию молока (но не его количество). В некоторых случаях у сук может наблюдаться ложная лактация, развивающаяся спонтанно, или как следствие кастрации, проведенной на стадии метэструса. К вероятным причинам данной патологии следует отнести повышение концентрации пролактина в конце метэструса (в отсутствие беременности) в связи с недостаточным угнетением его выработки из-за повышения концентрации прогестерона. Кроме того, можно предположить, что прогестерон уменьшает чувствительность молочных желез к пролактину, а лактация в конце метэструса обусловливается снижением концентрации прогестерона, вырабатываемого желтым телом. Вероятно, эта особенность объясняется происхождением собак от диких предков: при стайном проживании циклы доминирующих самок синхронизируются с циклами нерожавших самок, которым обычно поручается воспитание щенков.
Прерывание беременности воздействием на секрецию прогестерона
Экспериментальные исследования показывают, что первоначально желтые тела практически автономны и в течение 20 первых дней метэструса способны вырабатывать прогестерон без лютеотропной поддержки со стороны гипофиза. После чего требуется лютеотропное воздействие как ЛГ, так и пролактина, причем пролактин особенно необходим, начиная с 30 дня. Сходство между метэструсом у беременных и небеременных заставляет усомниться в том, что именно воздействие эмбрионов служит сигналом к продолжению секреции прогестерона у сук. Такая ситуация резко отличается от наблюдаемой у многих других домашних животных, у которых присутствие эмбрионов обеспечивает сохранение желтых тел и продолжение секреции прогестерона в течение по крайней мере первой половины беременности. Причем сохранение желтых тел обеспечивается в результате угнетения синтеза простагландинов в матке, и поэтому даже однократная инъекция простагландина вызывает прерывание беременности. У сук такой механизм отсутствует, это демонстрируется тем фактом, что гистерэктомия не оказывает влияния на продолжительность метэструса. Кроме того, желтые тела у собак относительно устойчивы к воздействию простагландинов. Однократная инъекция вызывает у собак временное снижение концентрации прогестерона, а для длительного и существенного уменьшения уровня прогестерона, необходимого для прерывания беременности, требуется повторное введение простагландинов. Как правило, такие инъекции переносятся очень плохо и сопровождаются рвотой и диареей.
Установление роли пролактина для поддержания желтых тел после 30 дня привело к многочисленным попыткам искусственно сократить продолжительность метэструса и индуцировать эструс назначением стимуляторов дофамина (бромокриптина, каберголина, метерголина), применение которых рассмотрено в 16 главе.
Роды
Снижение концентрации прогестерона перед родами наблюдается у большинства домашних животных и объясняется угнетением у беременной самки надпочечниковой секреции стероидных гормонов, что приводит к выработке эстрогена и простагландина эндометрием. У сук родам предшествует резкое падение концентрации прогестерона с 30 нмоль/л (10 нг/мл) до < 6 нмоль/л (< 2 нг/мл) в плазме крови; это, как сейчас принято считать, играет роль пускового механизма родов. Данный факт подтверждается и тем, что назначение прогестерона вызывает задержку родов, а препараты (например, эпостан), блокирующие его секрецию, - стимуляцию.
Резкое снижение концентрации прогестерона служит индикатором приближения родов. Концентрация > 6 нмоль/л (> 2 нг/мл) свидетельствует о том, что роды не начнутся в ближайшие 14 часов.
Эндокринные тесты на беременность
Как указано выше, пик концентрации прогестерона наблюдается приблизительно на 30 день, после чего она постепенно уменьшается, достигая к концу беременности 30 нмоль/л (10 нг/мл). Градиент снижения концентрации прогестерона после 30 дня у беременных сук практически такой же, как у небеременных. Понятно, что измерение концентрации этого гормона в целях диагностики беременности не имеет смысла.
Другими важными для беременности гормонами являются эстрогены. У многих видов, вероятно, в том числе и у собак, синтез эстрогенов происходит в трофобластах эмбрионов. Анализ мочи на содержание сульфата эстрона (основного метаболита эстрадиола), проведенный через 3 недели после вязки, т. е. в период имплантации, теоретически может использоваться для диагностики беременности, однако данный метод не нашел применения. На более поздних сроках в плазме крови определяется эстрадиол, но его концентрация не выше, чем в фазу метэструса, поэтому тесты на эстрадиол не применяются на этой стадии.
Содержание белков, таких, как фибриноген, в плазме крови во время так называемой протеиновой фазы повышается на 30–40 день беременности, причем данное явление не характерно для соответствующей стадии метэструса у небеременных сук. Повышение концентрации фибриногена в плазме измеряют с помощью иммунологических методов, а также с помощью существующего готового теста на беременность, хотя с распространением ультразвуковых методов необходимость в таком тесте постепенно снижается.
АНЭСТРУС
При наличии беременности данную фазу можно определить как интервал между родами и проэструсом, во время которого обычно наблюдается лактация. При отсутствии беременности анэструсом называют интервал между окончанием лютеиновой фазы и началом проэструса. На практике установить окончание лютеиновой фазы достаточно сложно по причине крайне низкого уровня концентрации прогестерона в этот период: обычно он не превышает 3 нмоль/л (1 нг/мл) к 90 дню после пика ЛГ (т. е. анэструс длится по крайней мере 60 дней до начала следующего проэструса). Концентрации эстрадиола и ЛГ изменяются в течение анэструса, однако эти изменения незначительны, и для их выявления требуются многократные анализы крови. Недавними исследованиями установлено, что, несмотря на наличие маленьких фолликулов в период анэструса, они не чувствительны ни к эндогенной, ни к экзогенной гормональной стимуляции. Вероятно, данное обстоятельство связано с рецепторами, но независимо от причины именно это является одним из факторов, вызывающих затруднения в индуцировании фертильного эструса у сук.
Относительно высокая концентрация эстрадиола отмечается в течение 10–20 дней до появления внешних проявлений проэструса, что свидетельствует о довольно продолжительном периоде выхода из анэструса. Механизм, запускающий изменения, вероятно, включает активизацию секреции ЛГ, наблюдаемую у некоторых других видов на соответствующей стадии репродуктивного цикла, - например, непосредственно перед переходом от анэструса к эструсу у видов, имеющих сезонный характер размножения, таких, как овцы. Брачный период овец связан с продолжительностью светового дня, обеспечивающего повышение активности гипофиза, и, следовательно, яичников, однако половой цикл домашней собаки не зависит от изменений продолжительности светового дня.
Изменения секреции пролактина могут послужить основанием для других объяснений анэструса у сук. Так, концентрация пролактина повышается в период метэструса и в начале анэструса, особенно при беременности. Пролактин обеспечивает лактацию в период анэструса у собак, как и у других видов (свиноматок, кобыл). Этот гормон вызывает снижение чувствительности яичников к гонадотропинам у других видов животных, однако у сук не выявлена последовательная временная корреляция между снижением концентрации пролактина и повышением уровня ЛГ в начале проэструса. При назначении агонистов дофамина (препаратов, подавляющих секрецию пролактина) на стадии метэструса или анэструса концентрация пролактина в плазме крови снижается, соответственно сокращается анэструс.
ЗАКЛЮЧЕНИЕ
В заключение следует отметить, что влияние гормонов на половой цикл домашней собаки почти такое же, как и у других домашних животных, а основное различие заключается в скорости перехода от одной фазы к другой. Для выявления характерных признаков этих фаз в главе приведены сведения об основных методиках, позволяющих ветеринарному врачу определить время наступления фертильного периода. Кроме того, в данной главе имеется информация, касающаяся процедур, позволяющих воздействовать на репродуктивный цикл суки. Знакомство с физиологией нормального цикла обеспечит понимание методов, применяемых для такого воздействия.
ЛИТЕРАТУРА
Concannon P. W., Morton D. B. and Weir B. J. (1989) Dog and cat reproduction, contraception and artificial insemination. Journal of Reproduction and Fertility, Supplement 39 .
Concannon P. W., England G. C. W., Verstegen J. P. and Russell H. A. (1993) Fertility and infertility in dogs cats and other carnivores. Journal of Reproduction and Fertility, Supplement 47 .
England G. C. W., Allen W. E. and Porter D. J. (1989) A comparison of radioimmunoassay with qualitative and quantitative enzyme-linked immunoassay for plasma progesterone detection in bitches. Veterinary Record 125 , 107–108.
Jeffcoate I. A. and Lindsay F. E. F.(1989) Ovulation detection and timing of insemination based on hormone concentrations, vaginal cytology and the endoscopic appearance of the vagina in domestic bitches. Journal of Reproduction and Fertility, Suppl. 39 , 277–287.
Lindsay F. E. F. (1983) The normal endoscopic appearance of the caudal reproductive tract of the cyclic and non-cyclic bitch: post-uterine endoscopy. Journal of Small Animal Practice 24 , 1–15.
ГЛАВА 2
Особенности физиологии и эндокринологии кошек
Дж. Верстеген (John Verstegen)
ВВЕДЕНИЕ
За последнее десятилетие была создана внушительная база данных, касающихся взаимосвязи поведенческих, яичниковых и эндокринных изменений, происходящих в организме кошек в течение анэструса, эструса и лютеиновой фазы после стерильной или фертильной вязки, во время беременности и в послеродовой период. Однако в знаниях о репродуктивной физиологии и эндокринологии все еще существуют пробелы и противоречия, требующие дальнейших исследований.
Эстральный цикл у кошек имеет сезонный характер (хотя бывают исключения) и во многом сходен с циклом таких млекопитающих, как кролики, норки и хорьки, особенно в том, что касается индукции овуляции.
ПОЛОВОЕ СОЗРЕВАНИЕ
Средний возраст полового созревания у домашней кошки варьирует. Как правило, первый эструс начинается, когда вес животного достигает 2,3–2,5 кг. В норме самка может достичь половой зрелости не раньше, чем в 4 месяца и не позднее, чем в 18 месяцев, но в большинстве случаев первые признаки сексуальной активности проявляются в 6–9 месяцев.
В какой-то степени возраст полового созревания определяется наследственностью, некоторые породы демонстрируют раннее или, наоборот, относительно позднее созревание. Наиболее "скороспелыми" являются короткошерстные породы (сиамская, бирманская), животные достигают зрелости при меньшем весе, чем кошки длинношерстных пород (персидская), у которых первая течка может начаться в возрасте 18 месяцев. Однако насколько можно судить, основным фактором, определяющим сроки полового созревания и установление цикла, являются длительность и интенсивность светового дня или искусственного освещения (для животных, содержащихся в помещении).
Достижение половой зрелости зависит от времени года и, как правило, происходит в то время, когда длительность светового дня увеличивается. Самки, вступающие в пубертатный период в конце лета, на фоне продолжительного светового дня демонстрируют поведенческий эструс в более раннем возрасте, чем те, что достигают зрелости осенью или зимой, - в последнем случае течка наблюдается лишь следующей весной.
Фиг. 2.1.
СЕЗОННЫЙ ХАРАКТЕР ЭСТРАЛЬНОГО ЦИКЛА
В отсутствие беременности или при ложной беременности эструс у кошек возобновляется каждые 2–3 недели весной, летом и осенью. При содержании в помещении с искусственным освещением, особенно если свет горит и ночью, периодичность цикла может проявляться вне зависимости от времени года, в таких случаях течка у кошек наблюдается и зимой. Влияние освещения на репродуктивный цикл осуществляется через эпифиз и вырабатываемый им гормон мелатонин. Механизм воздействия, видимо, сходен с тем, что наблюдается у кобыл.
Поведенческий цикл
С точки зрения поведения цикл кошки можно условно разделить на период течки и период ее отсутствия.
Проэструс и эструс
Период течки разделяют на проэструс и эструс. Поведенческие проявления проэструса и эструса наблюдаются в течение 3–10 дней. Переход от проэструса к эструсу у кошек в отличие от собак не выражен, поскольку за исключением поведенческих обе фазы не характеризуются заметными внешними проявлениями. Эструсом называют период, когда кошка испытывает половое возбуждение и подпускает к себе самца. Первые дни течки (с 1 по 4 день), на протяжении которых самка демонстрирует поведенческий эструс, но не подпускает к себе самца, называют проэструсом. Определить фазу проэструса с применением клинических методов затруднительно, поскольку некоторые самки подпускают к себе самца одновременно с наступлением поведенческого эструса, т. е. минуя подготовительный период проэструса. Развитие фолликулов в период проэструса начинается на фоне сниженной концентрации эстрадиола, недостаточной для обеспечения максимальных проявлений поведенческого эструса.
Начало эструса определяют по тому, что кошка подпускает к себе самца, а о его завершении судят по окончанию таких поведенческих проявлений. Период эструса характеризуется максимальным уровнем эстрогенов, вырабатываемых фолликулами.
В период течки кошка трется о людей и предметы, катается, зачастую проявляет крайнее дружелюбие. Характерным признаком являются протяжные призывные крики. При поглаживании, особенно по спине, кошка принимает характерную позу - выгибает поясницу, приподнимает круп, отводит в сторону хвост и перебирает лапами. Самцы проявляют интерес к самке, которая, вступив в период течки, может убежать из дома для случки. Во время вязки кошка издает громкие крики и с силой, а иногда и агрессивно пытается освободиться от самца, удерживающего ее за холку. После вязки половое возбуждение на некоторое время утихает; однако вскоре за первой следуют новые, многократные копуляции, как правило, наблюдаемые на протяжении всего эструса. Существует предположение, что вязки сокращают период эструса, хотя проверить справедливость этого утверждения в условиях контролируемого разведения достаточно сложно.
Осмотр наружных половых органов не позволяет выявить изменений, однозначно свидетельствующих о наступлении эструса. В некоторых случаях наблюдается небольшое покраснение или припухание вульвы, однако выделения отсутствуют. Иногда становится видна половая щель, незаметная в период анэструса.