ВитаминТ Е• + L• → LH + Витамин Е окисл
В результате ПОЛ происходит преобразование обычных липидов в первичные продукты ПОЛ (гидроперекиси липидов). Это приводит к появлению в мембранах участков ("дыр"), через которые наружу выходит содержимое как самих клеток, так и их органелл.
Первичные продукты ПОЛ разрушаются с образованием вторичных продуктов ПОЛ: альдегидов, кетонов, малонового диальдегида, диеновых коньюгатов. Накоплением в крови малонового диальдегида (МДА) объясняется синдром интоксикации, сопровождающий многие заболевания внутренних органов. Реагируя с SH- и СН3-группами белков, МДА подавляет активность цитохром-оксидаз (угнетая тем самым тканевое дыхание) и гидроксилаз. МДА обуславливает также ускоренное развитие атеросклероза.
При взаимодействии МДА с аминогруппами фосфолипидов образуются конечные продукты ПОЛ – Шиффовы основания. Примером этих соединений является пигмент липофусцин, появляющийся на оболочке глаза, на коже с возрастом. Липофусцин представляет собой смесь липидов и белков, связанных между собой поперечными ковалентными связями и денатурированными в результате взаимодействия с химически активными группами продуктов ПОЛ. Этот пигмент фагоцитируется, но не гидролизуется ферментами лизосом, накапливается в клетках, нарушая их функцию.
Негативные последствия активации ПОЛ:
1. Повреждение липидного бислоя мембран, в результате чего в клетки проникает вода, ионы натрия, кальция, что приводит к набуханию клеток, органелл и их разрушению.
2. Преждевременное старение клеток и организма в целом.
3. Взаимодействие высокореактивных продуктов ПОЛ с аминогруппами белков с образованием Шиффовых оснований.
4. Изменение текучести (вязкости) мембран, в результате чего нарушается транспортная функция мембран (функционирование ионных каналов).
5. Нарушение активности мембраносвязанных ферментов, рецепторов.
Активация ПОЛ характерна для многих заболеваний и патологических состояний:
1. атеросклероз и другие сердечнососудистого заболевания;
2. поражения ЦНС (болезнь Паркинсона, Альцгеймера);
3. воспалительные процессы любого генеза;
4. дистрофия мышц (болезнь Дюшенна);
5. онкологические заболевания;
6. радиационные поражения;
7. бронхолегочные патологии.
Антиоксидантные системы организма
В организме токсическое действие активных форм кислорода предотвращается за счет функционирования систем антиоксидантной защиты. В норме сохраняется равновесие между окислительными (прооксидантными) и антиоксидантными системами. Антиоксидантная система защиты представлена ферментными и неферментативными компонентами.
Ферменты антиоксидантной системы:
1. супероксиддисмутаза,
2. каталаза,
3. пероксидаза (глутатионпероксидаза),
4. глутатионредуктаза.
Наиболее активны эти ферменты в печени, почках и надпочечниках.
Супероксиддисмутаза превращает супероксидные анионы в пероксид водорода:
2•О2 + 2Н → Н2О2 + О2
Супероксидисмутаза является мощным ингибитором свободнорадикального окисления в организме, защищающим биополимеры (белки, нуклеиновые кислоты и др.) от окислительной деструкции. Супероксидисмутаза – индуцируемый фермент, т.е. синтез его увеличивается, если в клетках активируется ПОЛ.
Каталаза является гемопротеином и катализирует реакцию разложения пероксида водорода:
2Н2О2 → 2Н2О + О2
В клетках каталаза локализована в пероксисомах, где образуется наибольшее количество пероксида водорода, а также в лейкоцитах, где она защищает клетки от последствий "респираторного взрыва".
Глутатионпероксидаза – важнейший фермент, обеспечивающий инактивацию пероксида водорода и пероксидных радикалов. Он катализирует восстановление пероксидов при участии трипептида глутатиона. SH-группа глутатиона служит донором электронов и, окисляясь образует дисульфидную форму глутатиона:
Н2О2 + 2НS-глутатион → 2Н2О + глутатион-S-S-глутатион
Окисленный глутатион восстанавливается глутатионредуктазой:
глутатион-S-S-глутатион + НАДФН+Н → 2HS-глутатион + НАДФ
Глутатионпероксидаза в качестве кофермента использует селен. При его недостатке активность антиоксидантной защиты снижается.
Неферментативные антиоксиданты:
1. Природные водорастворимые антиоксиданты (витамин С; карнозин; таурин; восстановленные тиолы, содержащие SH-группы; цистеин; НS-КоА; белки, содержащие селен). Витамин С участвует в ингибировании ПОЛ с помощью двух механизмов. Во-первых, он восстанавливает окисленную форму витамина Е и поддерживает необходимую концентрацию этого антиоксиданта в мембранах клеток. Во-вторых, витамин С взаимодействует как восстановитель с водорастворимыми активными формами кислорода и инактивирует их.
2. Липофильные низкомолекулярные антиоксиданты, локализованные в мембранах клеток (витамин Е; β-каротин; КоQ; нафтахоиноны). Витамин Е – наиболее распространенный антиоксидант в природе, способен инактивировать свободные радикалы непосредственно в гидрофобном слое мембран и тем самым предотвращать развитие цепи перекисного окисления. b-каротин, предшественник витамина А, также ингибирует ПОЛ. Уменьшение содержания этого антиоксиданта в тканях приводит к тому, что продукты ПОЛ начинают производить вместо физиологического патологический эффект.
Растительная диета, обогащенная витаминами Е, С, каротиноидами, уменьшает риск развития атеросклероза и заболеваний сердечно-сосудистой системы, обладает антиканцерогенным действием. Действие этих витаминов связано с ингибированием ПОЛ и кислородных радикалов и, следовательно, с поддержанием нормальной структуры компонентов клеток.
Глава 12. Биохимия гормонов
Гормоны (от греческого hormaino – побуждаю) – это биологически активные вещества, которые выделяются эндокринными клетками в кровь или лимфу и регулируют в клетках-мишенях биохимические и физиологические процессы.
В настоящее время предложено расширить определение гормонов: гормоны – это специализированные межклеточные регуляторы рецепторного действия.
В этом определении слова "специализированные регуляторы" подчеркивают, что регуляторная – главная функция гормонов; слово "межклеточные" означает, что гормоны вырабатываются одними клетками и извне действуют на другие клетки; рецепторное действие – первый этап в эффектах любого гормона.
Биороль гормонов.
Гормоны регулируют многие жизненные процессы – метаболизма, функции клеток и органов, матричные синтезы (транскрипцию, трансляцию) и другие процессы, определяемые геномом (пролиферацию, рост, дифференцировку, адаптацию, клеточный шок, апоптоз и др.)
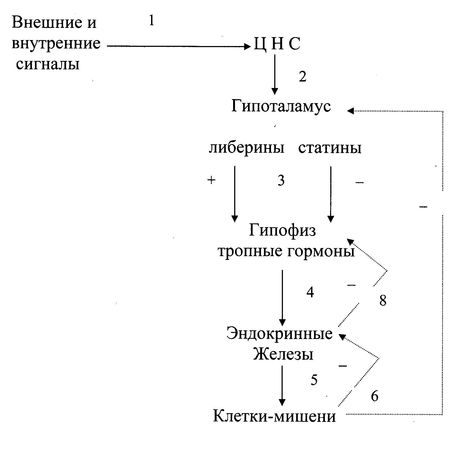
Рис. 12.1. Схема взаимосвязи регуляторных систем организма.
Эндокринная система функционирует в тесной взаимосвязи с нервной системой как нейроэндокринная.
1. Синтез и секреция гормонов стимулируются внешними и внутренними сигналами, поступающими в ЦНС.
2–3. Эти сигналы по нейронам поступают в гипоталамус, где стимулируют синтез пептидных рилизинг-гормонов (либеринов и статинов), которые стимулируют или ингибируют синтез и секрецию гормонов передней доли гипофиза.
4–5. Гормоны передней доли гипофиза (тропные гормоны) стимулируют образование и секрецию гормонов периферических эндокринных желез, которые поступают в кровь и взаимодействуют с клетками-мишенями.
Уровень гормонов в крови поддерживается благодаря механизмам саморегуляции (регуляция по принципу обратной связи). Изменение концентрации метаболитов в клетках-мишенях подавляет синтез гормонов в эндокринной железе или в гипоталамусе (6, 7). Синтез и секреция тропных гормонов подавляется гормонами эндокринных желез (8).
Классификация гормонов
Гормоны классифицируются по химическому строению, биологическим функциям, месту образования и механизму действия.
Классификация по химическому строению.
По химическому строению гормоны делят на 3 группы (табл. 12.1):
1. пептидные или белковые;
2. производные аминокислот;
3. стероидные
4. производные арахидоновой кислоты – эйкозаноиды (оказывают местное действие)
Классификация гормонов по химическому строению
Пептидные (белковые)
1. Кортикотропин
2. Соматотропин
3. Тиреотропин
4. Пролактин
5. Лютропин
6. Лютеинеизирующий гормон
7. Фолликулостимули-рующий гормон
8. Мелоноцитстимули-рующий гормон
9. Вазопрессин
10. Окситоцин
11. Паратгормон
12. Кальцитонин
13. Инсулин
14. Глюкагон
Производные аминокислот
1. Адреналин
2. Норадреналин
3. Трийодтиронин (Т3)
4. Тироксин (Т4)
Стероиды
1. Глюкокортикоиды
2. Минералокорти-коиды
3. Андрогены
4. Эстрогены
5. Прогестины
6. Кальцитриол
Клетки некоторых органов, не относящихся к железам внутренней секреции (клетки ЖКТ, клетки почек, эндотелия и др.), также выделяют гормоноподобные вещества (эйкозаноиды), которые действуют в местах их образования.
Классификация гормонов по биологическим функциям
По биологическим функциям гормоны можно разделить на несколько групп (табл. 12.2.)
Таблица 12.2. Классификация гормонов по биологическим функциям.
| Регулируемые процессы | Гормоны |
|---|---|
| Обмен углеводов, липидов, аминокислот. | Инсулин, глюкагон, адреналин, кортизол,тироксин,соматотропин. |
| Водно-солевой обмен. | Альдостерон, вазопрессин. |
| Обмен кальция и фосфатов. | Паратгормон, кальцитонин, кальцитриол. |
| Репродуктивная функция. | Эстрогены, андрогены, гонадотропные гормоны. |
| Синтез и секреция гормонов эндокринных желез. | Тропные гормоны гипофиза, либерины и статины гипоталамуса. |
Эта классификация условна, поскольку одни и те же гормоны могут выполнять разные функции. Например, адреналин участвует в регуляции обмена липидов и углеводов и, кроме этого, регулирует артериальное давление, частоту сердечных сокращений, сокращение гладких мышц. Эстрогены регулируют не только репродуктивную функцию, но и оказывают влияние на обмен липидов, индуцируют синтез факторов свертывания крови.
Классификация по месту образования
По месту образования гормоны делятся на гормоны:
1. гипоталамуса
2. гипофиза
3. щитовидной железы
4. паращитовидных желез
5. поджелудочной железы
6. надпочечников
7. половых желез.
Классификация по механизму действия
По механизму действия гормоны можно разделить на 3 группы:
1. Гормоны, не проникающие в клетку и взаимодействующие с мембранными рецепторами (пептидные, белковые гормоны, адреналин). Сигнал передается внутрь клетки с помощью внутриклеточных посредников (вторичные мессенджеры). Основной конечный эффект – изменение активности ферментов;
2. гормоны, проникающие в клетку (стероидные гормоны, тиреоидные гормоны). Их рецепторы находятся внутри клеток. Основной конечный эффект – изменение количества белков-ферментов через экспрессию генов;
3. гормоны мембранного действия (инсулин, тиреоидные гормоны). Гормон является аллостерическим эффектором транспортных систем мембран. Связывание гормона с мембранным рецептором приводит к изменению проводимости ионных каналов мембраны.
Основные свойства и особенности действия гормонов
1. Высокая биологическая активность. Гормоны регулируют метаболизм в очень малых концентрациях – 10–8 – 10–11М.
2. Дистантность действия. Гормоны синтезируются в эндокринных железах, а биологические эффекты оказывают в других тканях-мишенях.
3. Обратимость действия. Обеспечивается адекватным ситуации дозированным освобождением и последующими механизмами инактивации гормонов. Время действия гормонов различно:
• пептидные гормоны: сек – мин;
• белковые гормоны: мин – часы;
• стероидные гормоны: часы;
• йодтиронины: сутки.
4. Специфичность биологического действия.
5. Плейотропность (многообразие) действия. Например, катехоламины рассматривались как краткосрочные гормоны стресса. Затем было выявлено, что они участвуют в регуляции матричных синтезов и процессов, определяемых геномом: памяти, обучения, роста, деления, дифференциации клеток.
6. Дуализм регуляций (двойственность). Так, адреналин как суживает, так и расширяет сосуды. Йодтиронины в больших дозах увеличивают катаболизм белков, в малых – стимулируют анаболизм.
Рецепторы гормонов
Биологическое действие гормонов проявляется через их взаимодействие с рецепторами клеток-мишеней. Клетки, наиболее чувствительные к влиянию определенного гормона, называют клеткой-мишенью. Специфичность гормонов по отношению к клеткам-мишеням обусловлена наличием у клеток специфических рецепторов, которые входят в состав плазматических мембран.
Рецепторы – это специфические структуры клетки, обладающие высоким сродством по отношению к одному определенному гормону.
Рецепторы по химической природе являются, сложными белками (гликопротеинами). Рецепторы пептидных гормонов и адреналина располагаются на поверхности мембраны и содержат три домена. Первый домен расположен на внешней стороне клеточной мембраны, содержит гликозилированные участки и обеспечивает узнавание и связывание гормона. Второй домен – трансмембранный. Третий (цитоплазматический) домен создает химический сигнал в клетке.
Рецепторы стероидных и тиреоидных гормонов содержат три функциональные области:
1. домен узнавания и связывания гормона;
2. домен связывания с ДНК;
3. домен, отвечающий за связывание с другими белками, вместе с которыми участвует в регуляции транскрипции.
Механизм передачи гормональных сигналов через мембранные рецепторы
Гормоны (первичные посредники) связываются с рецепторами на поверхности клеточной мембраны и образуют комплекс гормон-рецептор. Этот комплекс трансформирует сигнал первичного посредника путем изменения концентрации внутри клетки вторичных посредников. Вторичными посредниками являются: циклический АМФ (цАМФ), цГМФ, инозитолтрифосфат (ИФ3), диацилглицерол (ДАГ); Са, NO (оксид азота II).
Аденилатциклазная система.
Гормоны, взаимодействие которых с рецептором клетки-мишени приводит к образованию цАМФ действуют через систему, включающую: белок-рецептор, G-белок и фермент аденилатциклазу.
Известно более 200 различных G-белков. В отсутствие гормона G-белок связан с ГДФ и неактивен. Образование комплекса гормон-рецептор приводит к конформационным изменениям G-белка, замене ГДФ на ГТФ и активации G-белка. Существуют GS-стимулирующий и GI-ингибирующий аденилатциклазу белки.
Последовательность событий, приводящих к изменению активности аденилатциклазы:
1. связывание гормона с рецептором;
2. комплекс гормон-рецептор взаимодействует с G-белком, изменяя его конформацию;
3. вследствие изменения конформации G-белка происходит замена ГДФ на ГТФ;
4. комплекс GS-белок•ГТФ активирует аденилатциклазу (комплекс GI-белок•ГТФ ингибирует аденилатциклазу);
5. активация аденилатциклазы приводит к увеличению скорости образования цАМФ из АТФ.
Далее образовавшийся под действием аденилатциклазы цАМФ активирует протеинкиназу А. Активированная протеинкиназа А фосфорилирует ферменты и другие белки, что сопровождается изменением функциональной активности белков-ферментов (активацией или ингибированием).
Протеинкиназа – это внутриклеточный фермент, который может существовать в двух формах. В отсутствие цАМФ протеинкиназа представлена тетрамером, состоящим из двух каталитических (2С) и двух регуляторных (2R) субъединиц (неактивный фермент). В присутствии цАМФ протеинкиназный комплекс обратимо диссоциирует на одну 2R-субъединицу и две свободные каталитические субъединицы С. Субъединицы С обладают ферментативной активностью.
Гуанилатциклазная система.
Эта система, генерирующая цГМФ как вторичный посредник, сопряжена с гуанилатциклазой. Этот фермент катализирует реакцию образования цГМФ из ГТФ (подобно аденилатциклазе). Молекулы цГМФ могут активировать транспортные системы мембран клеток или активируют цГМФ-зависимую протеинкиназу G, которая участвует в фосфорилировании других белков в клетке.
Циклические нуклеотиды запускают каскады реакций аденилатциклазного или гуанилатциклазного механизмов регуляции активности ферментов. Одна молекула гормона, активирующая рецептор, может "включать" несколько G-белков. Каждый из них в свою очередь активирует несколько молекул аденилатциклазы с образованием тысяч молекул цАМФ или цГМФ. Образующийся вторичный посредник усиливает сигнал в тысячу раз. Суммарное усиление сигнала равно 106 – 107 раз.
Снятие гормонального сигнала достигается уменьшением концентрации вторичного посредника. Реакции превращения цАМФ или цГМФ в неактивные метаболиты АМФ или ГМФ катализируют ферменты фосфодиэстеразы.