Место не каждой холостяковой залежки может обеспечить условия (на берегу и вокруг), необходимые для выживания приплода, и соответственно лишь некоторые из них могут превратиться в продуцирующие лежбища. Но движение в этом направлении неизбежно.
Итак, в пределах акватории, известной обитателям исходного разбухшего лежбища, появляется одно или несколько сателлитных лежбищ, гаремные территории которых будут еще более притягательны для переселенцев, чем холостяковые залежки на их месте. И отток, и приток особей обладают некоторой инерцией. Так что молодые лежбища еще некоторое время будут расти ускоренно за счет переселенцев и собственной продукции, пока сначала переселенцы, а потом и часть собственных потомков не будут блокированы разбуханием территории уже дочернего лежбища. Из-за той же инерции отток с родительского лежбища не прекратится сразу, как будут выравнены продукция и выселение, и лежбище не только остановится в росте, но и начнет сокращаться в размерах, либо сжимаясь к центру, либо распадаясь на несколько самостоятельных кусков.
Уменьшение размеров неизбежно позднее пресечет выселение, и начнется вначале очень медленный рост за счет только своего небольшого поголовья. Цикл замкнулся.
От освоения нового необжитого места под холостяковую залежку, на которой спустя десять-двадцать лет появятся первые беременные самки и еще лет через десять начнется медленный рост гаремной территории, через фазу ускоряющегося роста к продуцированию выселенцев и образованию удаленных холостяковых лежек (будущих дочерних лежбищ), а затем и к обособлению от событий на прочих лежбищах в пределах собственной резко сократившейся территории должно проходить не меньше полувека, если не век.
В этой упрощенной схеме не учтены по крайней мере три группы факторов, влияющих на смену и длительность фаз: ограниченность пригодных мест для береговых лежбищ (кусок пригодного берега может быть длиной и сто метров, и в несколько километров) и кормовых ресурсов в море; включение регулятора - детской смертности при повышении плотности населения и влияние выборочного (или невыборочного) промысла на структуру данного лежбища. Но сам ход событий как будто не противоречит известному об отариидных лежбищах сейчас.
Универсальны ли пространственные структуры
Рассмотрев структуру отариидной колонии, возможности многовидовых конгломератных поселений и историю отдельной колонии от детства к зрелости, попробуем примерить отариидную схему к на первый взгляд похожей второй половине рис. 21, где Е. В. Сыроечковский нарисовал часть гусиной колонии.
Вся наличная площадь ее поделена между отдельными владельцами - гусаками, изгоняющими все постороннее, каждый со своего участка. На колонии остаются только территориальные самцы и взрослые самки, участвующие в размножении в этом сезоне. Если вычленить основной элемент конструкции колонии, то обнаружатся прочные многолетние пары, в которых место для гнезда выбирает гусыня, а привязанный к ней на невидимый поводок гусак старается не отстать и одновременно очистить ближайшую округу радиусом от одного до пятнадцати метров (в зависимости от обстоятельств радиус меняется) от попадающих туда гусей одного с ним ранга, т. е. от любого гусака. Невольно вспомнишь секача, преследующего на залежке среди разнополого и разновозрастного зверя избранную самку и очищающего свое индивидуальное пространство вокруг от конкурентов.
К месту гнездования весной гуси приходят уже парами, сбившимися в стаи по нескольку десятков особей, вместе с некоторыми родителями держатся их прошлогодние птенцы (числом от 1 до 5). Уже через неделю после появления первых гусей на острове место будущей колонии распределено между отдельными парами, рассеянными с интервалами от нескольких до нескольких десятков метров. Взаимное расположение гнезд определяется притягательностью места колонии для многих гусынь и невозможностью для привязанных к ним гусаков располагаться близко друг к другу. Место выбирает самка по своим неясным для нас меркам, с учетом качеств биотопа и привязанности к местам своих прошлых гнезд.
После выбора точки на местности самка, а следовательно, и самец с индивидуальным обороняемым пространством вокруг него прочно закреплены в ней. Различные интервалы между гнездами могут быть связаны как с непригодностью отдельных участков, неотличимых на первый взгляд от соседних, занятых, так и с личными качествами гусаков. Закрепленный на местности гусак как будто ничем не отличается от территориального секача. Оба они будут защищать свою землю от посторонних и пытаться спариваться с любой попавшей туда самкой своего вида. То, что гусыня вместе с гусаком нападает на вторженцев, показывает, что и у нее в это время есть свое охраняемое пространство. В то же время гусак, по крайней мере к концу насиживания, покидая надолго гнездо с гусыней, как бы выходит на это время из своего индивидуального пространства, и ничто не показывает наличие неприкосновенной зоны вокруг него (опять же полная аналогия секачу, покинувшему на время свое место на гаремном лежбище).
Таким образом сформировавшаяся колония представляет собой местами плотное, местами диффузное скопление закрепленных на местности индивидуальных пространств гусаков. Прошлогодние птенцы и не участвующие в этом году в гнездовании одиночки и не закрепившиеся пары активно изгоняются резидентами (в том числе и собственными родителями) и в разгар сезона уже кочуют группами по острову.
Отклонения от схемы двояки: гусак, потерявший гусыню или ушедший от нее достаточно далеко, пытается спариваться с любой гусыней, независимо от того, в паре она или нет; часть гусынь в парах либо по реальной непригодности незанятых другими участков, либо по индивидуальным особенностям поведения не находит места для гнезда и откладывает яйца в чужие гнезда или около них.
За гусыней, прорывающейся к чужому гнезду, следует ее гусак, что неизбежно ведет к конфликту с хозяином, который то гонит вторженцев, то пытается спариваться с ними независимо от их половой принадлежности. В результате таких попыток некоторая часть территориальных гусаков успевает за сезон покрыть по нескольку прошедших через их индивидуальное пространство самок, т. е. проигрывает нормальный для территориальных секачей ушастых тюленей вариант.
Первыми колония отторгает прошлогодков, изгнанных из родительских индивидуальных пространств. Затем уже в тундре, вдали от колонии, к ним присоединяются не участвовавшие в размножении или потерявшие кладку (или партнера) особи, державшиеся до этого в пределах колонии. Птицы сбиваются в крупные (до 700 птиц) табуны, где индивидуальные пространства особей, если и сохраняются, то радиус их минимален. У гусаков на гнездовье индивидуальные пространства прикреплены к гнездам до вылупления гусят. Якорь, державший территорию самца на месте, оказывается поднятым, и гусак уходит из колонии, не отставая от гусыни с гусятами и отгоняя вторженцев от своей передвижной территории.
Выводки один за другим покидают гнездовье. Уже двух-, трехнедельные гусята участвуют в защите от вторженцев (сначала от мелких, потом и от более крупных) общего для семьи индивидуального пространства гусака. Попыток раздела пространства внутри семьи не видно. К подъему выводка на крыло индивидуальное пространство оказывается сжавшимся до предела, как уже случилось раньше у незагнездившихся особей.
Сначала вылинявшие годовики, потом взрослые неудачники и, наконец, вылинявшие выводки начинают группами покидать остров, двигаясь к местам зимовок. При этом места зимовок разных пар одной колонии могут оказаться существенно различными. Как обстоит дело с индивидуальным пространством особей на зимовках, неясно. Но именно на зимовках происходит образование новых пар, а также иногда восстановление старых прерванных связей. Это и определяет поведение гусака - носителя индивидуального пространства - на следующий год или часто на многие годы (т. е. репродуктивное перемешивание происходит на местах зимовок и затруднено на местах гнездовий). В то же время на зимовке сохраняются еще связи родителей с последним выводком.
Образование конгломератных колоний с совместным гнездованием разных видов гусей не редко и связано, помимо привлекательности одних и тех же угодий для разных гусей, еще и с отчетливым предпочтением компании (даже других видов) одиночеству и малочисленности. При этом дистанция между гнездами разноплеменников меньше, чем между гнездами единоплеменников, т. е. индивидуальное пространство гусака, закрытое для своих, частично открыто для чужих.
Наиболее занятным результатом влечения гусиных пар к иноплеменникам, да еще очень далеким, пожалуй, будет гнездование в пределах активно охраняемого индивидуального пространства гнездящейся пары белых сов (или других крупных хищников). Исследовавший это рискованное сожительство Е. В. Сыроечковский полагает, что именно оно позволяет первым десяткам пар вселиться и окрепнуть в месте, защищенном присутствием гнезда хищников от прочих охотников до гусиных яиц, положив тем самым начало новой растущей колонии.
Вероятно, начало гусиной колонии может быть и другим (например, на островке или в другом защищенном месте), но несомненно, что начало кладут не одиночки, а немногочисленные пары, по каким-то причинам порвавшие со своим местом рождения и нуждающиеся в годах спокойного гнездования на новом месте, при постоянном приращении площади гнездовья и сохранении плотной упаковки индивидуальных пространств гусаков. Лишь после этого спокойного роста за счет туземного приплода и переселенцев из других мест будет сформирован достаточно прочный костяк из достаточного количества достаточно прочно закрепленных и достаточно скученных гнезд, который может противостоять разрушительному действию непогоды и хищников.
Выйдя из-под опеки и продолжая расти, гнездовье должно пройти, по-видимому, те же фазы, что и отариидная колония, с неодинаковостью работы разных зон колонии, неизбежным (при отсутствии помех) продуцированием выселенцев, усиленным оттоком их в новые места гнездования, сокращением территории гнездовья и временной стабилизацией перед повторением цикла. Разумеется, и здесь ограничение ресурсов (кормовых и территориальных), изменение смертности приплода и выборочное (или без разбора) разрушение гнезд будет определять ход и длительность отдельных фаз.
Индивидуальное пространство
Сравнение гусиной и отариидной колонии показывает, что в основе образования обеих лежит стремление большого количества особей закрепиться на ограниченном участке и претензии взрослых самцов не допускать в ближнее пространство вокруг себя (индивидуальное пространство) особей своего ранга. Эта мерка - недоступное для других пространство - позволяет довольно надежно описывать геометрию взаимного размещения особей в сообществе, ту основу, на которой из отдельных особей одного вида возникает постройка следующего порядка. Кроме того, введение этой мерки позволяет заменить ей одной многочисленные частные характеристики, прижившиеся в зоологической литературе, где отдельно называют дистанцию, активно поддерживаемую между особями в стаде (индивидуальная дистанция), отдельно - охраняемое вокруг движущейся особи пространство (передвижное пространство), отдельно - охраняемое пространство вокруг оседлой особи (территория), хотя все три - лишь характеристики единого механизма, препятствующего сбиванию в кучу. Индивидуальное пространство либо перемещается вместе с владельцем, либо оказывается закрепленным в какой-то точке местности. Кроме того, варьируют величина владения, его проницаемость и степень скученности владений. То есть для достаточно полного описания можно обойтись ограниченным количеством параметров, комбинация которых даст любую конкретную конструкцию поселения.
Радиус индивидуального пространства и у гусаков, и у ушастых тюленей не превышает десятка длин тела, хотя обычно гораздо меньше (1–2 длины). Он может меняться в каких-то пределах у одной и той же особи, позволяя, например, после ухода соседей по колонии оставшимся несколько расширить свои владения, так же как и гусаку в момент перехода его гусыни к оседлости (до этого момента радиус не превышает 10 м, после - может достигнуть 16 м).
Внутрь индивидуального пространства самца и у гусей, и у морских котов допускаются лишь самки и сеголетки, но у морских львов к ним добавляются и субординатные самцы более старших возрастов. Основным контингентом владельцев индивидуального пространства и у гусей, и у отариид стали взрослые самцы в сезон размножения. Но его можно найти и у самок (гусыня защищает пространство вокруг кладки и выводка, но только в присутствии гусака, а котиха при благоприятных условиях долго защищает пространство вокруг щенка в одиночку), и у детенышей (у щенков - в играх со сверстниками, а у гусят - при защите родительского индивидуального пространства от гусят других выводков).
Необходимость смыкать края индивидуальных пространств соседей можно предполагать с большей уверенностью для морских котов, с меньшей - для морских львов и гусей (но и здесь одиночное гнездование пары - скорее результат утраты прежде бывших соседей).
И у гусей, и у отариид четко наблюдается фантастичная на первый взгляд ситуация присутствия индивидуального пространства в отсутствие владельца, если он перед уходом успел оставить достаточно следов в памяти соседей.
Напротив, владелец может обнаруживать свое индивидуальное пространство либо только в присутствия определенной особи, за которой и следует неотрывно (гусак с еще незагнездившейся гусыней, секач с одиночной самкой вне гаремного лежбища, котиха со щенком), либо даже в отсутствие полового партнера (секач до прихода самок, ищущий место на будущем гаремном лежбище). А кроме того, возможны два внешне похожих варианта: 1) владелец пространства привязан только к точке на местности (гусыня у еще пустой гнездовой ямки и территориальный секач на лежбище) и 2) владелец привязан к особи, закрепленной на местности (гусак при загнездившейся гусыне, гусыня у кладки, котиха у только что родившегося и неспособного к перемещениям щенка).
Можно еще больше упростить описание и построить таблицу, расположив по горизонтали характеристики индивидуального пространства, а по вертикали - способы ограничения перемещения владельца, и, получив в ее клетках возможные варианты, сопоставить их с реализуемыми в наблюдаемой модели.
Из сравнения же гусиной и котовой моделей обнаруживается не только нехватка прямых наблюдений (событий вне сезона размножения; индивидуальной, возрастной и сезонной изменчивости поведения), но и возможность увидеть некоторые частные и более общие закономерности.
Так, для понимания возникновения гнездового паразитизма у птиц (наиболее известна обыкновенная кукушка) важна наблюдаемая в гусиной колонии ситуация, когда часть пар оказывается незакрепленной на местности и гусыни либо бросают свои яйца поблизости от чужих гнезд, либо с риском для жизни из-за активной обороны хозяевами своего индивидуального пространства откладывают яйца в чужие гнезда, т. е. практикуют гнездовой паразитизм внутри вида. Неясно, насколько это определяется качествами гусыни и насколько - конкретной ситуацией в колонии.
Несколько иную окраску получает и такая, казалось бы, простая проблема, как отличие полигама от моногама. Гусака-моногама и секача-полигама различают не столько количество оплодотворенных за сезон самок (через территорию гусака нередко проходит несколько гусынь, а многие секачи, долго просидевшие на гаремном лежбище, так и не увидели самок на своих территориях), сколько привязанность владельца индивидуального пространства к конкретной самке или конкретному месту.
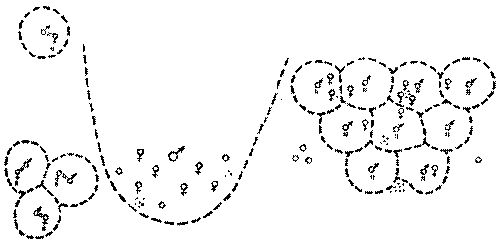
Рис. 22. Три схематизированные модели пространственного размещения особей
Слева - белые гуси, посредине - калифорнийские морские львы, справа - морские коты. Самки и территориальные самцы обозначены обычными для полов значками, субординатный самец - кружком, детеныши - точками, границы индивидуального владения очерчены прерывистой линией, а привязанность особи к другой особи или точке на местности показана короткими парными штрихами. Обращает внимание различный способ ограничения перемещений особи в гусиной и котовой моделях, а также большие размеры и неотчетливость части пограничной линии в индивидуальном владении калифорнийского морского льва
Но главное, что у таких далеких таксономически и биологически, как сравниваемые, просматривается общая тенденция к уравновешиванию скучивания многих особей на ограниченном участке проявлением у некоторых групп населения (здесь у взрослых самцов) владения индивидуальными пространствами. Набор вариантов индивидуального пространства и способов взаимодействия их владельцев достаточно ограничен, чтобы поддаваться классификации (рис. 22). Особи, в норме не обнаруживающие наличия индивидуального пространства (здесь самки и молодняк), сохраняют потенциальную способность к его проявлению, а владельцы в разных ситуациях демонстрируют разные варианты индивидуального пространства.
Так же и в пределах обоих таксонов потенциально возможны разные варианты индивидуальных пространств, т. е. набор их общ, а употребляемость различных вариантов неодинакова и норма для одного таксона оказывается редкостью в другом, В результате при явном несходстве способов ограничения подвижности носителей индивидуального пространства можно найти у морских котов типичные гусиные варианты (секач на холостяковой лежке, привязанный к избранной самке и неотвязно следующий за ней), а у гусей - типично котовые, когда через закрепленное на местности индивидуальное пространство гусака проходят свободно перемещающиеся по местности самки (хоть и в сопровождении собственных гусаков).
Обе колонии сходны и тем, что собраны из индивидуальных пространств взрослых самцов и недоступны для всех, кроме полового партнера владельца и последнего приплода. Различие же моделей - в противоположных способах закрепления владельцев индивидуальных пространств на местности, разной степени их скученности (индивидуальные пространства смыкаются краями у котов и более дисперсны у гусей) и в большей отчетливости намеков на владение индивидуальными пространствами у гусят и гусынь по сравнению с котовыми щенками и котихами.
Итак, на двух половинках рисунка 21 - похожие постройки, но сделанные из разного строительного материала. Набор стройматериалов небезграничен, и, так же как пропорции черепа ленивца повторяют пропорции черепа ушастого тюленя, так и строение котового лежбища оказывается и в целом, и в деталях повторяющим скопление собравшихся для размножения североамериканских лягушек-быков, с разделением участка между территориальными самцами с последующим прохождением через их сплоченные индивидуальные пространства позднее приходящих готовых к икрометанию самок.
Сравнение колоний разных групп животных помогает увидеть общие для всех законы и специфику каждого варианта, но не восполняет недостаток прямых наблюдений. А даже для наиболее изученных северных морских котов неизвестно, где и как включаются в структуру лежбища впервые размножающиеся самцы и самки, чем определяется интенсивность обмена особями внутри лежбища и между соседними лежбищами, как влияет более чем вековой выборочный выбой молодых самцов на окраинах лежбищ на расселение зверей и структуру лежбищ. Без этих знаний приходится постоянно сталкиваться с несовпадением реального положения в стаде с прогнозами.