Вершинные участки подводных поднятий, как правило, имеют скальное дно, полностью лишенное рыхлых осадков, что само по себе определяет доминирование рыб-планктофагов в сообществах таких биотопов. До самого недавнего времени бытовали два взгляда на источник пищевых ресурсов, определяющих существование и продукцию "горных" популяций рыб. Согласно одному из них, продуктивность поднятий определяется локальным орографическим эффектом - подъемом глубинных вод, обусловленным завихрением набегающего на гору потока постоянного течения и приводящим в действие механизм биологической продуктивности в его полном цикле: от повышения концентрации биогенных солей в эвфотическом слое к увеличению первичной продукции водорослей и за ее счет к увеличению биомассы зоопланктона. Для более точного ("более наукообразного") объяснения этого явления была выдвинута гипотеза о формировании над вершинами квазистационарных вихрей, которые и обеспечивают постоянный подъем вод, будучи настолько изолированными от окружающих вод, что внутри них образуются самостоятельные экологические системы. Действительно, подводный рельеф, усложняя в известной степени структуру набегающего потока, может способствовать образованию локальных вихрей, приводящих в конечном счете к обогащению поверхностных слоев биогенными элементами. Однако пространственно-временная устойчивость таких вихрей представляется довольно слабой, а их продолжительная изоляция - в высшей степени проблематичной. Ведь время, необходимое для развития фитопланктона, исчисляется несколькими сутками, а кормовая база рыб - мезопланктон и макропланктон - развивается намного дольше, соответственно около месяца и полугода. Столь длительное существование над подводной возвышенностью неподвижных и существенно замкнутых вихревых образований почти невероятно: если влияние локального обогащения поверхностных вод биогенами и может отразиться на продуктивности зоопланктона и тем более на рыбопродуктивности, то произойдет это только в значительном удалении от места подъема вод. Таким образом, эффект увеличения биомассы кормовых организмов за счет локального "апвеллинга" может быть использован не бентопелагическими рыбами той подводной возвышенности, которая послужила причиной возникновения такого "шлейфового" пятна повышенной биомассы планктона, а только истинно пелагическими хищниками (например, тунцами и кальмарами). Поэтому гораздо более правомочным представляется другой подход, связывающий кормность поднятий с горизонтальным приносом (адвекцией) пищевых объектов набегающим потоком. Прямые исследования питания рыб, живущих над подводными возвышенностями, показывают, что их откорм происходит в основном за счет макропланктона, входящего в состав мигрирующих звукорассеивающих слоев (миктофиды, эвфаузииды и т. п.) и опускающегося днем на самую вершину горы (если, конечно, она лежит выше нормальной глубины дневного опускания этих слоев).
Распространение. Географическое распространение океанических рыб, входящих в состав бентопелагического комплекса, еще не изучено в деталях, но представляется, что размеры их ареалов сильно варьируют. Некоторые из них весьма обширны - такие виды, как, например, эпимезобентальные антигония Antigonia capros и рыба-бекас Macrorhamphosus scolopax, мезобентальные мягкоголовый долгохвост Malacocephalus laevis и низкотелый берикс Beryx splendens, мезобатибентальные альдровандия Aldrovandia phalacra и антимора Antimora roslrata, батибентальные гладкоголов нарцет Narcetes stomias и ксиелациба Xyelacyba myersi, батиабиссобентальные гладкоголов беллоция Bellocia koefoedi и батизавр Bathysaurus mollis, абиссобентальные слепой тифлонус Typhlonus nasus и эхиномакрурус Echinomacrurus mollis, обнаружены на соответствующих глубинах в тропических водах всех трех океанов. В то же время во всех вертикальных зонах есть и виды, которые известны из очень ограниченных участков и, по-видимому, на самом деле имеют очень узкое распространение (хотя представления об эндемизме могут основываться на недостаточных данных). Крайний случай почти "точечного" ареала демонстрирует распространение латимерии, обитающей только на склонах двух маленьких островов Коморского архипелага. Большинство других узколокальных эндемиков также обитает на вершинах подводных поднятий.
Известно, что дробность биогеографического деления бентали, достигающая наибольшей выраженности на шельфе и в верхней части склона (т. е. на глубинах залегания поверхностных и подповерхностных вод), уменьшается в нижней батиали и сводится к минимуму на ложе океана, где можно обнаружить только самые главные границы (те, которые разделяют регионы высшего ранга - области). Многими работами показано, что главные зоогеографические границы в мезобентали как бы продолжают вглубь границы, проходящие на шельфе. Действительно, анализ географического распространения рыб, населяющих склоны континентов и подводные поднятия, дает возможность выявить те же крупномасштабные регионы, что и в сублиторальной зоне. Так, Г. А. Головань, рассматривая ареалы мезобентальных рыб, обитающих у западных побережий Африки и Европы, обнаружил в составе ихтиофауны 12 географических элементов и выделил четыре района - бореальный, северный субтропический, тропический и южный субтропический - в районировании шельфа.
Немногочисленность поимок многих видов батибентальных рыб сильно затрудняет анализ их географического распространения. Тем не менее представления о преобладании в этой зоне узколокальных ареалов, существовавшие до недавнего времени, не подкрепляются, как правило, новыми данными. Почти для всех видов, известных по многим нахождениям, характерно широкое распространение. Достаточно четкое разделение существует, однако, между фаунами тропических, умеренных и холодных вод. В тропической зоне выделяются экваториальный и широкотропический типы ареалов с циркумглобальным, индо-вестпацифическим, атлантическим и восточно-тихоокеанским подтипами. Умеренноводные ареалы подразделяются на потальные и бореальные, а холодноводные - на антарктические и арктические.
Что касается абиссобентальных и хадальных рыб, все они известны по немногим, часто единичным находкам, что сильно затрудняет рассмотрение их зоогеографии. Нужно отметить, впрочем, что на ложе океана некоторые виды уже сейчас найдены в далеко разобщенных районах (в двух или трех океанах встречены Bathymicrops regis, Ipnops meadi, Holcomycteronus profundissiraus, Abyssobrotula galatheae и др.), и это позволяет предполагать их широкое распространение.
Глава 5. Происхождение океанических рыб
Общепризнано, что жизнь на Земле зародилась в прибрежной зоне океана. Там же, на мелководьях, возникли все типы и почти все классы свободноживущих животных (об этом не раз писал академик Л. А. Зенкевич), в том числе все современные классы рыбообразных и рыб – миксины, миноги, хрящевые и костные. Именно оттуда шло заселение пресных вод и суши, а также, вполне естественно, всех биотопов "открытого океана", рассматриваемых в этой книге. Меня всегда занимал вопрос, каким образом происходило это заселение. Как возникли столь существенные различия между населением донно-придонных и пелагических биотопов? Почему одни из океанических видов почти не отличаются от родственных прибрежных форм, другие хотя и имеют специфические черты, но легко выводятся от мелководных родственников, третьи относятся к аберрантным группам, положение которых в системе отнюдь не очевидно? Есть ли соответствия между степенью систематической обособленности рыб, населяющих разные биотопы открытого океана, древностью этих рыб и очередностью заселения ими тех или иных биотопов? Нужно добавить еще, что палеонтологических свидетельств эволюции океанических рыб практически нет, и ответы на все возникающие вопросы могут быть получены только на основе косвенных доказательств.
Впервые я коснулся этих проблем в своей книге "Ихтиофауна океанской эпипелагиали", но вовсе не все, сказанное 20 лет назад, представляется мне верным сейчас, и в первую очередь это касается заселения глубоководной пелагиали, которое, как я думал тогда, шло из верхних горизонтов водной толщи.
Представления об эволюционной истории и расселении любой фауны зиждутся па изученности ее систематического состава, так как естественная система организмов должна, как известно, отражать их филогению. В отношении рыб и рыбообразных такая система, впрочем, еще не разработана, и их классификация, даже на уровне самых высоких категорий, активно обсуждается и перестраивается в последние годы. "Классические" представления, синтезом которых была общепринятая вплоть до середины 60-х годов система академика Л. С. Берга, претерпели очень существенные изменения. В этой книге используется в основном классификация, принятая в недавней сводке Дж. Нельсона (1984) и дополненная данными более поздних исследований*.
* Число отрядов щележаберных, в соответствии с трудами Л. Компаньо, увеличено с 5 до 12, из отрядов карпозубообразных (Cyprinodontiformes) и бериксообразных выделены отряды сарганообразных и китовидкообразных, отряды иглообразных (Syngnathiformes), индостомообразных (Indostomiformes) и пегасообразных (Pegasiformes) включены в отряд колюшкообразных, а отряд долгоперообразных (Daclylopteriformes) - в отряд скорпенообразных.
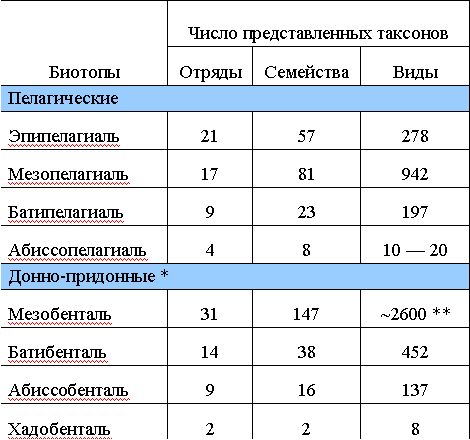
Сопоставление систематического состава прибрежных (шельфо-неритических) и океанических таксоценов показывает значительные различия между ними. Как видно из табл. 5, в прибрежной зоне представлено 42 из 44 отрядов рыб и рыбообразных, встречающихся в соленой воде (в это число включены и такие отряды, в которых есть хотя бы отдельные диадромные виды, проводящие часть жизни в море), и только два отряда - стомиеобразные и миктофообразные в полном составе специфичны для открытого океана. В океанской пелагиали живут виды из 24 отрядов, в бентали и бентопелагиали - из 31 отряда, но далеко не все они могут считаться процветающими вдали от берегов.
Из круглоротых, например, миксины очень характерны для мезобентали, тогда как миноги (они размножаются только в пресной воде) играют ничтожную роль как в этом биотопе (трехзубая минога Entosphenus tridentatus встречается, впрочем, в северной Пацифике вплоть до глубины 1000 м), так и в эпипелагиали (Geotria australis отмечена вдали от берегов в субантарктических водах). Современные цельноголовые (кроме хоботнорылых химер рода Callorhynchus) явно тяготеют к мезобентали, хотя ископаемые формы, как полагают, были приурочены к шельфовым морям. Хорошо представлены в открытом океане щележаберные рыбы - лишь два из 12 отрядов не выходят за пределы прибрежной зоны. Заселение океанических биотопов акулами и скатами также шло главным образом вниз по склону. В мезобентали и мезобентопелагиали представлено 23 семейства из 9 отрядов, в батибентали и батибентопелагиали - 3 семейства из 3 отрядов, причем 5 семейств встречаются только на глубинах, а еще 3 имеют там центр своего разнообразия. Пелагические океанические виды существуют в 5 отрядах щележаберных (не считая гнюсообразных, случайно проникающих в открытые воды) и 9 семействах (опять же без ксеноэпипелагическпх); два семейства акул полностью ограничены в своем распространении эпимезопелагиалью, а одно (Lamnidae) представлено за ее пределами только нерито-океаническим кархародоном. Из числа костных рыб в открытом океане живут виды 22 отрядов, в том числе 19 отрядов (более 140 семейств) в донно-придонных биотопах и 17 отрядов (94 семейства) в пелагиали.
Таблица 5
Систематический состав (число семейств) морских прибрежных и океанических ихтиоценов

В целом ихтиофауна открытого океана образует значительную но объему и хорошо обособленную часть мировой фауны рыб (табл. 6). Она включает, по самым предварительным подсчетам (для более точных оценок время еще не наступило), около 1100 видов, живущих в толще воды без какой-либо связи с дном и около 2800 видов бентопелагического комплекса (без талассосублиторальных видов, число которых, по самой грубой прикидке, составляет не менее 300 - 400), в том числе почти 1700 видов, не выходящих на шельф. В полном объеме океаническая ихтиофауна содержит, таким образом, около 3900 (2200 без шельфо-батиальных форм) видов (см. табл. 2 - 4), т. е. составляет более четверти всех морских рыб (прибрежных рыб насчитывается около 9,5 тыс. видов). Несколько отличающиеся оценочные цифры приводит Д. Коэн, по подсчету которого число видов прибрежных рыб достигает 9100, а океанических - 2500 (в том числе 255 эпипелагических, 1280 глубоководных донно-придонных и 1010 глубоководных пелагических).
Таксономическая обособленность отдельных биотопических группировок океанических рыб довольно сильно различается. Для эпипелагиали, например, специфичны всего 16 семейств рыб (30% от числа представленных в биотопе), причем большая их часть характерна для ее нижних слоев, пограничных с мезопелагиалыо. Полностью океаническим является, строго говоря, только одно из верхнеэпипелагических семейств - макрелещуковые. Специфичность на родовом и видовом уровнях достигает тем не менее 50% списочного состава.
Глубоководные пелагические ихтиоцены значительно более своеобразны. Их самобытный облик определяется главным образом наличием ряда характерных таксонов высокого ранга, таких, как отряды стомиеобразных, миктофообразных, китовидкообразных, подотряд глубоководных удильщиков и др., очень разнообразных в видовом отношении. Все они отсутствуют в прибрежной зоне и очень слабо представлены в толще воды над материковым склоном. Половина мезобатипелагических семейств (42 из 83), большинство родов (61%) и почти все виды (85%) также специфичны для этих биотопов.
Таблица 6
Систематическое разнообразие океанических ихтиоценов
(число представленных таксонов)
* Весь бентопелагический комплекс (бенталь + бентопелагиаль).
** Без эпимезобентальных около 1500 видов.
Придонные ихтиоцены открытого океана представляются менее обособленными, но и они имеют характерные отряды (целакантообразные, спиношипообразные) и подотряды (например, гладкоголововидные). Число специфических семейств относительно невелико - всего 20% (27 из 137), хотя на уровне родов и видов эти таксоцены вполне четко отличаются от прибрежных и пелагических, а также друг от друга.
В целом наиболее резко выражены границы между совокупностями пелагических и донных таксоценов рыб, причем степень выраженности этих границ явно увеличивается с глубиной. В вертикальной последовательности бентальных и бентопелагических ихтиоценов самая четкая разделительная полоса соответствует краю континентальной отмели, но, как указывалось в предыдущей главе, многие преимущественно шельфовые виды все же опускаются в верхние участки материкового склона. Дальнейшие изменения таксономического состава - от мезобентали к батибентали и далее к абиссобентали и хадали - происходят постепенно и наиболее заметны на видовом уровне. В толще воды границы между эпи-, мезо- и батипелагиалью, несмотря на существование зон смешения (экотонов), выражены более четко, чем границы между гомологичными донно-придонными биотопами.
Исходным материалом для формирования рецентной фауны океанических рыб бесспорно были шельфовые и неритические предковые формы. Из этого постулата со всей определенностью следует, что именно прибрежные участки (или по меньшей мере некоторые из них) действовали в определенные периоды времени, а быть может, действуют и сейчас как эволюционные центры, продуцирующие виды, способные к заселению новых биотопов за пределами этих участков, т. е. к переходу в новую среду обитания*. Очевидно также, что прямо из шельфо-неритической зоны могли заселяться только непосредственно прилегающие к ней биотопы, которые, в свою очередь, обеспечивали заселение контактирующих с ними участков среды. Последовательность появления вселенцев в том или ином биотопе может в первом приближении определяться их ранжированием по систематической обособленности: наличие видов, общих с исходным (донорским) биотопом, говорит о совсем недавнем или еще не закончившемся вселении, тогда как дивергенция на видовом и родовом уровне, а тем более на уровне семейств, подотрядов и отрядов свидетельствует о все увеличивающемся сроке, прошедшем со времени успешной колонизации.
* Альтернативой такому представлению могло бы служить только явно абсурдное в данном аспекте предположение о некогда сплошном распространении предковых форм в относительно однородной среде и последующем расчленении их ареалов в результате образования физических барьеров (так называемая "викариация" в понимании сторонников кладизма).
Основываясь на этих представлениях, рассмотрим теперь, как происходило заселение донно-придонных океанических биотопов. В мезобентали очень обычны рыбы, характерные и для шельфовой зоны (см. табл. 4). Большинство этих эврибатных эпимезопелагических видов входит в состав экотонного сообщества кромки шельфа и самой верхней батиали и не опускается глубже 400 - 500 м. Среди них, несомненно, есть и такие формы, которые потенциально способны к адаптивной модификации, дающей возможность закрепиться на глубинах материкового склона.
Примеры такого закрепления показывают мезобентальные виды преимущественно шельфовых родов, например Saurida kaianus (сем. Synodontidae) или Chaetodon guyotensis (Chaetodontidae). Есть в этом биотопе и специфические роды, принадлежащие к преимущественно шельфовым семействам - Ostichthys (сем. Holocentridae), Aphanopus и Assurger (Trichiuridae), Bembradium (Platycephalidae) и др. К эндемичным мезобентальным семействам относятся плащеносные акулы и акулы-домовые, ложноромбовые и шестижаберные скаты из щележаберных, глубоководные солнечники, хаунаксовые и холлардиевые из костистых рыб. Только в мезобентали сохранился до нашего времени: и единственный представитель кистеперых рыб - латимерия.