Имеется зона регулярных контактов и потоков фуражиров. Это гнездовой вал. Частота контактов невысока и сводится к периодическому обмену тактильными сигналами*, а также к постоянному наблюдению за ситуацией в ближайшей части соседнего сектора гнездового вала. Сильное возбуждение муравьев в пограничной зоне одного из секторов привлекает туда часть фуражиров из соседней колонны. При этом отдельные фуражиры могут увлечься и попасть в чужой поток фуражиров. Такие переходы возможны только на гнездовом валу или в непосредственной близости от него. На пространственной обособленности колонн на кормовом участке все это не сказывается: потоки продолжают отталкиваться друг от друга, а сам участок остается поделенным между колоннами. Основанное на общей бдительности взаимодействие фуражиров помогает муравьям заблаговременно выявить угрозу гнезду. Надо отметить, что это всегда реакция отдельных особей или небольших групп муравьев. Чтобы активизировалась колонна в целом, источники возбуждения должны действовать непосредственно в ее гнездовом секторе. Действия колонн при этом не согласуются. Единственным местом на поверхности гнезда, воздействие на которое может вызвать общую ответную реакцию семьи, является маковка купола.
Следует отметить, что схемы взаимодействия муравьев перечисленных выше функциональных групп различны. Во внутреннем конусе каждая колонна обменивается молодью непосредственно со всеми остальными колоннами. Чтобы установить это, мы провели следующий опыт. Куколок с разноцветными метками выложили в несколько приемов на разные кормовые дороги гнезда.
Муравьи тут же начали заносить куколок в гнездо. Почти одновременно меченые куколки начали поступать в соседнее родственное гнездо, куколки с каждой новой меткой составляли определенную часть от общего числа выложенных на соответствующей дороге.
Обмен по принципу "каждая с каждой" отражает и поддерживает определенный тип взаимодействия колонн в семье муравьев. Здесь действуют многосторонние связи без какого-либо предпочтения кому-либо из партнеров.
Возможность многостороннего контакта имеется и у особей, занятых на поверхности купола. Правда, не у всех, а лишь у тех муравьев, которые функционируют на самой маковке, где смыкаются границы всех секторов. Здесь обычно максимальное число наблюдателей, так как это самое ответственное место муравейника, маковку, восстанавливают в первую очередь. Эта часть гнезда оформляется сразу же, даже если место, над которым оно должно находиться, оказалось в результате разрушения на уровне почвы.
Возможно, в какой-то мере маковка воплощает целостность муравейника.
Действительно, разделить верхнюю часть купола на две, сделать его двуглавым - значит положить начало глубокому и долгому процессу перестройки всей организации семьи. Нередко это приводит даже к разделению муравейника на два.
На остальной поверхности купола колонны контактируют и обмениваются сигнальной информацией только по эстафете. Преднамеренной передачи информации из колонны в колонну нет; муравьи воспринимают как сигналы тревоги специфические позы других особей в минуты опасности. Не имеет значения, принял ли такую позу муравей из своей или соседней колонны. Состояние возбуждения поддерживается специальными веществами-феромонами тревоги, или торибонами, выделяемыми муравьями при появлении угрозы.
Эстафетные контакты фуражиров разных колонн на гнездовом валу связаны с ролью взаимодействия особей данной функциональной группы в жизни семьи. В результате такого взаимодействия устанавливаются радиальные структуры кормового участка, а для сохранения целостности семьи контакты фуражиров значения не имеют.
Вообще гнездовой вал играет у многих видов муравьев особую роль. Здесь организационная структура семьи трансформируется в пространственную структуру кормового участка и начинается раздел сфер влияния колонн вне гнезда: определяются направления кормовых дорог или потоков фуражиров в тех случаях, когда постоянных дорог нет. Дальнейшее размежевание колонн на территории осуществляется при взаимодействии отталкивающихся друг от друга потоков фуражиров. В остальных частях кормового участка границы поддерживаются группами фуражиров, численность которых в пограничных зонах обычно выше, нежели в среднем по участку.
В итоге складывается достаточно сложная и вместе с тем рациональная система постоянного взаимодействия колонн в гнезде и на территории, схематично изображенная на рис. 8. И хотя колонны обособлены и во многом развиваются почти независимо друг от друга, они все-таки составляют единое целое - семью.
Подытоживая изложенное в этом разделе, следует назвать два фактора, наиболее важных для сохранения семьи муравьев: общее гнездо и регулярные обмены особями Гнездо важно не только как жилище, общая крыша. В таком гнезде присутствует и постоянно выравнивается единый запах семьи. Ведь именно по запаху муравьи отличают своих от чужих. Пока муравейники имеют одинаковые или близкие запахи, воины между ними не будет. Встретившись на куполе или вдали от него рабочие муравьи первым делом обмениваются легкими ударами антенн. В булавах антенн находятся хеморецепторы, с помощью которых муравей и определяет запах различных предметов, в том числе и запах муравьев. Свой будет отпущен или привлечен к какой-нибудь деятельности. Чужому не поздоровится; его либо разорвут на части, либо изгонят.
Обмены как средство сохранения лояльных отношений колонн дополняют общность гнезда и общность гнездового запаха.
Однако обмены оказываются крайне необходимыми при выполнении еще одного определяющего условия устойчивости семьи: сравнимости колонн по численности.
Сравнимость колонн по численности населения
Сколько-нибудь выраженной иерархии колонн в пределах семьи нет. В секционных гнездах это обусловлено тем, что все секции единого гнезда равны между собой. Одинаковы по численности и колонны, населяющие секции. Это показано на разных видах, относящихся к различным родам и подсемействам.
Численность населения одной секции стабильна для каждого вида в конкретных условиях обитания. А поскольку отношения группировок муравьев одного вида отражают в конечном счете соотношение численности этих группировок, то основа для доминирования какой-либо из колонн в семьях, обитающих в секционных гнездах, отсутствует.
У стабилизированной семьи рыжих лесных муравьев все обстоит сложнее, так как с сооружением купола численность муравьев перестает зависеть от емкости подземной части гнезда. Купол можно надстроить, увеличив его диаметр и высоту. Увеличился объем купола - возросли возможности гнезда вместить дополнительное количество особей. Поэтому какой-то определенной численности колонны здесь нет.
В гнезде одного размера может быть разное число колонн. Так, у северного лесного муравья в гнезде с диаметром купола 1 м оказывается и две, и пять колонн. Естественно, что в первом случае численность каждой колонны выше. Но внутри семьи колонны, как правило, соизмеримы. И чем старше муравейник, чем меньше вокруг других гнезд, так или иначе влияющих на его рост, тем ровнее длина дорог, интенсивность движения на них и общая численность колонн. В идеальной ситуации, когда одиночный муравейник расположен в равномерном по составу, возрасту и продуктивности насаждении, кормовой участок приобретает форму круга.
Такие семьи обладают наибольшей устойчивостью во времени, и только серьезное вмешательство извне может вывести их из стабилизированного состояния. В этом случае чем больше колонн в муравейнике, тем меньше вероятность последующих серьезных изменений в структуре семьи. И все же такие изменения происходят. Гнездо растет, и у него появляются новые дороги. На некотором расстоянии от старого муравейника возникают новые муравейники, которые тоже начинают расти. Наступает время, и здесь образуются колонны.
Что же приводит к этим изменениям? Ведь очень часто новые колонны возникают задолго до того момента, когда уже имеющиеся достигли верхнего предела численности. Такой предел еще не известен, однако в совершенно одинаковых условиях по соседству существуют муравейники с разной численностью колонн. Например, у каждого по три колонны, но колонны первого гнезда насчитывают по 50-60 тыс. особей, второго - 90-100 тыс., а третьего - 150-200 тыс. особей. Можно было бы ожидать, что новая колонна возникнет прежде всего в последнем, наиболее мощном муравейнике. Однако в действительности очень часто дело обстоит иначе. Следовательно, абсолютная численность еще не определяет ситуации.
Чтобы выяснить, в чем причина, был проведен следующий опыт. В 1969 г. мы подобрали шесть близких по размерам муравейников обыкновенного лесного муравья, имевших по три равных между собой дороги. Территория каждого подопытного муравейника была тщательно закартирована до начала опыта. Все шесть муравейников были одиночными. Четырем из них на одну из дорог в течение июля-августа выкладывали в больших количествах куколок того же вида, взятых из гнезд, не участвовавших в опыте. В несколько приемов в каждом случае на подопытную дорогу было выложено по 100 тыс. куколок.
Вышедшие из куколок рабочие муравьи влились в состав соответствующей колонны, которая таким образом резко усилилась. Два других муравейника в это же время также были усилены куколками, но одновременно все колонны усиливались на 90-100 тыс. особей каждая.
Реакция муравейников в первом и втором вариантах оказалась различной. Равномерное усиление всех колонн привело к росту гнезд и также равномерному увеличению дорожной сети каждой колонны. Усиление одной из колонн нарушило имевшееся до опыта равновесие. Кормовой участок такой колонны резко расширился, дорога удлинилась и появились новые ответвления. Контрольные дороги не изменились. Наконец, одна из усиленных колонн разделилась и выделила дочернюю семью*. Отводок* был сооружен в сентябре того же года, через месяц после усиления колонны куколками.
Из приведенного опыта следует несколько выводов. Обмены, которые производят муравьи внутри гнезда, не обеспечивают выравнивания численности колонн в случае резкого усиления одной из них. Если одна из колонн вследствие каких-либо причин начинает расти быстрее остальных, различия в численности накапливаются. В определенный момент подобная несоразмерность может послужить причиной выделения отводка наиболее сильной колонной.
Интересно, что в различных экспериментах и при естественном ходе событий дочернюю семью всегда выделяет самая мощная колонна, резко выделяющаяся по численности особей. Есть все основания расценивать образование отводка как средство сохранения структуры семьи: проблема внутрисемейной регуляции решается путем расселения и создания новой семьи.
Образование отводка - один из путей выравнивания колонн по численности населения, наиболее радикальный, но не единственный. Второй, более обычный путь - формирование новой колонны. Здесь тоже обязательным условием является нарушение численного равновесия колонн.
Образование новой колонны теснее связано со структурой кормового участка, с дорожной сетью муравейника. Возникновение колонны у рыжих лесных муравьев невозможно без появления новой дороги. Число разветвлений у той или иной дороги роли не играет. Важно не число ответвлений на территории, а число потоков, выходящих из гнездового купола. Поэтому процесс разделения колонны начинается всегда на кормовом участке, схематично он изображен на рис. 9. Из схемы видно, что разделение в большей степени зависит от возможности радикального раздвоения исходного кормового участка колонны. Чем шире занимаемый участком сектор, тем проще использующей его колонне разделиться на две. Число колонн и связанных с ними дорог у рыжих лесных муравьев весьма постоянно. Еще в 30-х годах появились работы немецкого энтомолога Р. Штегера, проследившего топографическое постоянство кормовых дорог в течение 13 лет. Имеются подобные данные и у других авторов, работавших с муравьями разных родов. Смещения отдельных ответвлений дорог обычно не связаны с изменением в составе колонн. А иногда разделение колонны на две почти не отражается на периферийных отрезках дорог (именно этот вариант показан на рис. 9, в).
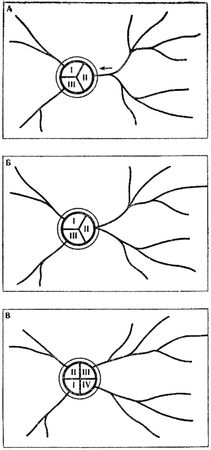
Рис. 9. Процесс образования новой колонны при раздвоении дороги
А - наметившееся разделение наиболее сильной и разветвленной дороги (стрелка показывает направление перемещения развилки);
Б - положение накануне разделения колонны на две: развилка переместилась к основанию купола;
В - колонна разделилась, ответвления стали самостоятельными колоннами
I, II, III, IV - номера колонн (дорог, секторов)
Увеличение колонн - процесс, направленный как бы внутрь муравейника, ведущий к умножению связей в семье, укрепляющий ее как единое образование. А это означает, что данный процесс в какой-то мере противодействует почкованию* - выделению дочерних муравейников. Действительно, чем больше колонн, тем прочнее каждая из них связана с семьей, тем труднее одной из них достичь условий, необходимых для выделения отводков.
Наибольшее значение имеют при этом два показателя: размер семьи, тесно связанный с размерами муравейника и кормового участка, и число колонн. В результате взаимодействия двух тенденций вырабатывается оптимальная зона образования отводков. Причем эта зона даже не видоспецифична вообще, а отражает специфику вида лишь в определенных условиях. Так, в подмосковном ельнике у обыкновенного лесного муравья максимум образованных отводков падает на муравейники с диаметром 1,2-1,3 м, состоящие из четырех колонн. А в сосняках у того же вида диаметр наиболее плодовитого гнезда - 1-1,1 м при трех колоннах.
Процессы разделения колонн и почкования противодействуют друг другу и имеют разные последствия для роста гнезда. Увеличение числа колонн - это путь к развитию крупных муравейников, появлению "сверхгнезд". Муравейники-гиганты почти всегда одиночные и всегда включают много колонн. Максимальное число колонн в средней полосе отмечено нами в Приокско-террасном заповеднике; семья малого лесного муравья имела 12 дорог. Охраняемая ею территория занимала более 1,5 га. Почкование, наоборот, сдерживает рост муравейника. Причем сдерживание происходит за счет самой мощной, т. е. наиболее быстро растущей колонны, что особенно важно. В итоге семья удерживается в пределах, соответствующих состоянию интенсивного выделения отводков.
Попробуем охарактеризовать этот размерный интервал с позиции оптимизации размеров семьи. Представим себе требования, которым должен соответствовать такой муравейник.
В общих чертах оптимальной организацией любого биологического объекта можно считать такую организацию, которая обеспечивает устойчивую плотность поселения на максимальном для данных условий обитания уровне. Это почти в равной мере применимо к различным уровням организации. Для муравьев мы вправе использовать такой подход по отношению к отдельной семье, комплексу муравейников* одного вида и даже к обитающим на одной территории поселениям муравьев разных видов, или, как их назвал в 1975 г. московский мирмеколог А. В. Демченко, к многовидовым ассоциациям муравейников*.
Для отдельного муравейника основным показателем оптимальности организации будет стабильность размеров семьи и гнезда во времени. Важны также вероятность гибели муравейника определенного размера, его возможности к дальнейшему росту и образованию отводков. Необходимо выбрать отрезок времени, соответствующий нашей задаче. Год для этой цели слишком малый срок: условия разных лет могут резко отличаться друг от друга, а это неизбежно сказывается на состоянии каждой семьи. Но пятилетнего периода уже достаточно, чтобы выявить общие тенденции развития муравейников. Вот что дал анализ пятилетних данных по 112 семьям северного лесного муравья, обитающего в ельнике-кисличнике в районе Солнечногорска Московской области.
Совсем маленькие муравейники (диаметр до 40 см), как правило, обречены. Если в течение двух лет такой муравейник не вырастет в более крупный, он погибнет. Погибает 83% подобных гнезд. Муравейники этого размерного класса не обладают устойчивостью. Среди муравейников покрупнее (диаметр 45-60 см) доля погибших меньше (24% за пять лет), но и здесь семья всегда перед альтернативой: либо вырасти, либо погибнуть. Лишь 4% семей сохраняется в исходном классе в течение пяти лет. Зато большинство муравейников (72%) вырастают, причем некоторые за это время достигают метра в диаметре.
Далее вероятность погибнуть уменьшается, но скорость роста начинает снижаться. Одновременно появляется и обратная тенденция: муравейники, продержавшись некоторое время в более высоком классе, вновь попадают в разряд более мелких. Это особенно заметно на примере крупных гнезд диаметром 1,25 м и более. Тенденции к росту резко падают с увеличением контрольного периода с одного до пяти лет. Такой муравейник может выйти в следующий класс на один-три года, но в течение пяти лет ни один из них не удерживается там и снова уменьшается в размерах, нередко даже ниже исходного уровня. Среди гнезд диаметром более 1,4 м в своем размерном классе удерживаются только из муравейников, а остальные - начинают деградировать.
Единственной устойчивой размерной группой оказались муравейники с диаметром купола 1,05-1,2 м. Большинство муравейников этого размера сохраняются в классе независимо от срока наблюдения. У них еще сильна тенденция к росту: почти 1/3 гнезд за пятилетие переходит в следующий размерный класс. Это значит, что муравейники такого размера, обретая устойчивость, сохраняют активность. Вероятность гибели муравейников данного класса за пятилетний период равна нулю. И лишь 7% гнезд уменьшается в размерах.
Но, пожалуй, самое интересное заключается в том, что муравейники именно этого диаметра образуют основную часть естественных отводков в комплексе. Следовательно, есть все основания рассматривать образование отводков как механизм поддержания численности семьи на определенном, оптимальном для данного вида в данных условиях уровне.
Что же это за уровень? Расчеты показали, что в гнезде северного лесного муравья диаметром 112,5 см (середина класса) обитает около 870 тыс. муравьев. Эти муравьи организованы, как правило, в четыре колонны (среднее число колонн - 4,11).