Знакомясь с двумя механизмами стабилизации муравейников, мы не должны упускать из виду одну важную общую черту. Действие этих механизмов непостоянно. Они срабатывают периодически, как ответная реакция семьи на накопившиеся количественные диспропорции между составляющими ее колоннами. И образование отводка, и формирование новой колонны - специфические аварийные меры сохранения семьи. И хотя фактологическая сторона процессов достаточно прояснилась, один из наиболее интригующих, узловых моментов остается неясным: какие реальные силы заставляют самую могучую колонну в семье выделять отводок? Почему именно так решается этот важный для муравейника вопрос? Ведь заглядывая немного вперед, можно сказать, что образование отводков далеко не всегда выгодно материнскому муравейнику*. Казалось бы, усилившаяся колонна могла бы потеснить соседей по гнезду или просто расширить свой сектор.
К сожалению, ответов на эти вопросы еще нет. Очевидно только, что существенного передела секторов гнезда в обычных условиях не происходит, так же никогда не наблюдаются внутригнездовые муравьиные войны.
В природе мы можем застать самые различные этапы оптимизации муравейника. Этот процесс складывается из двух процессов: оптимизации размеров семьи и оптимизации ее организации. Они взаимосвязаны, дополняют друг друга, но не совпадают. Синхронности здесь тоже нет, потому что первый процесс почти непрерывен, а второй - четко дискретен. Поэтому первый процесс обычно идет быстрее. Численность колонн "догоняет" его скачками (и даже может опередить).
При этом увеличение числа колонн связано с формированием наиболее рациональной формы кормового участка - округлой.
Формированию кругового кормового участка могут препятствовать различные факторы. Такой участок маловероятен при неравноценности кормовых угодий вокруг муравейника. Распределение растений с колониями насекомых, выделяющих падь, также имеет значение. Особенно сильно сказывается размещение соседних муравейников. Конечно, не всяких, а того же или конкурирующих видов. Установив в 1932 г. наличие у муравьев охраняемой территории, английский эколог Ч. Элтон положил начало целому направлению в изучении жизни муравьев - первых насекомых, у которых данный вопрос был разносторонне изучен.
Конкуренция сводится к борьбе за территорию, реже - за места поселения. Кормовые участки расположенных поблизости муравейников вытянуты в противоположные стороны либо параллельными полосами. Нередко форма их причудлива: кормовые дороги с прилегающими полосками угодий как бы пробираются среди чужих земель. В таких условиях выравнять колонны путем увеличения их числа нельзя. И тогда выручают отводки. Вместе же оба процесса дают большинству семей достаточно богатые возможности для достижения устойчивого состояния.
Колонна-муравейник в муравейнике
Достигая определенных размеров, семья муравьев начинает дробиться на части, каждая из которых вполне автономна и в состоянии полностью или почти полностью выполнять все необходимые для полноценного существования функции. Под одной крышей сосуществуют несколько вполне сформировавшихся муравейников? - Именно так и обстоит дело. Еще раз перечислим основные характеристики, указывающие на самостоятельность колонн. Сюда относятся пространственная и пищевая обособленность в гнезде и на кормовом участке, устойчивость состава, отсутствие передачи из колонны в колонну оперативной информации о состоянии и делах на территории, фактически автономное развитие.
У полигинных видов каждая колонна в состоянии перейти в самостоятельное гнездо. Это довольно часто происходит при сильном разрушении муравейников (например, кабанами). На месте разрушенного гнезда, на сохранившемся гнездовом валу или неподалеку от него, возникают несколько муравейников. Число таких гнезд обычно равно числу колонн в исходной семье. Иногда из семьи уходит только одна колонна, нередко часть колонн объединяется в одном из вторичных муравейников, а часть - в другом. При таких внутренних разделах соблюдается в общем-то лишь одно правило: гнезда меняются, семьи меняются, а колонны остаются, сохраняя свой кормовой участок и дорогу.
Но силы, ранее удерживавшие их в одном гнезде, сохраняются. По крайней мере первое время. Впрочем, понятие "первое время" здесь относительно, оно может растянуться и на несколько лет. Бывают и быстротечные варианты, когда уже через две-три недели все отношения между бывшими сородичами прекращаются. Это случается чаще при обосновании каждой колонной своего гнезда. Связи между групповыми вторичными муравейниками гораздо устойчивее.
Дальнейшая судьба вторичных муравейников различна. Колонны, прервавшие связи, становятся самостоятельными семьями и далее развиваются как одиночные муравейники. Если связи между гнездами устойчивы, события принимают почти фатальный характер. Вторичные гнезда исходно равны или почти равны друг другу, так как равны основавшие их колонны. Некоторое время, не более одного-двух лет, это равенство сохраняется. Между муравейниками существует оживленная дорога с интенсивным взаимным обменом особями. Обмен всегда равный - один к одному. Семьи продолжают придерживаться принципа, действовавшего, когда они еще были колоннами одного муравейника.
Но колонны развиваются с разной скоростью. И семьи тоже. Один муравейник растет немного быстрее, другой - медленнее или вообще не растет. Спустя некоторое время различия между ними становятся настолько существенными, что меньший становится чем-то вроде отводка у более крупного. И тогда более крупный поглощает, присоединяет его. Население маленького гнезда переселяется при этом в большое. Но опять же колонны остаются. Этапы такого воссоединения в одном гнезде схематично изображены на рис. 10. Отметим, что все это возможно только у относительно небольших одинарных муравейников. Если муравейники стали взрослыми и в них произошло разделение колонн, подобная ситуация уже исключена.
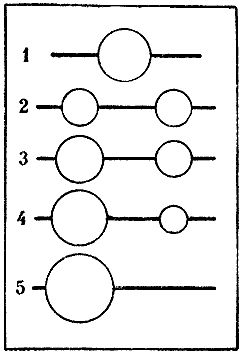
Рис. 10. Воссоединение колонн, перешедших в отдельные гнезда после разрушения исходного муравейника
1 - исходный муравейник с двумя колоннами,
2 - равные по размерам гнезда колонн непосредственно после обособления,
3 - нарушение равенства колонн в результате их неравномерного роста,
4 - различия в силе колонн достигают критической величины,
5 - колонны объединились в более крупном муравейнике, вслед за этим часто следует деление усилившейся колонны.
На первый взгляд объединение, в общем гнезде при таких обстоятельствах противоречит изложенному выше принципу выравнивания колонн путем выделения отводка. Там выделение, а здесь та же неравноценность колонн ведет к объединению. Однако противоречия нет. Отводок должна выделить наиболее сильная колонна, входящая в муравейник. Но если колонна в муравейнике одна, ей не с кем сравнивать свою мощь. Система из двух одноколонных муравейников после усиления одного из них начинает несколько напоминать материнский муравейник и отводок. Но и это лишь внешнее подобие, так как отводок может возникнуть только у муравейника с несколькими колоннами, т. е. ситуация искусственная и именно поэтому неустойчивая.
И все-таки система существует один-два года?! В том-то и дело, что неустойчивой система становится лишь после нарушения равновесия разбежавшихся по отдельным гнездам колонн. Когда различия в размерах семей становятся кратными, поглощение меньшей происходит быстро, как правило во время весеннего или осеннего массового обмена между гнездами. Такие обмены нередко интенсивнее летних. Поэтому они и являются обычно как бы поворотным пунктом в жизни муравейников.
В объединении муравейников проявляется очень сильная тенденция муравьев к концентрации. Об этом мы уже говорили в первой главе. Она даже доминирует при отсутствии взрослых муравейников, например, после искусственного переселения отводков. Аналогичная ситуация складывается и в результате распадения семьи на несколько мелких. Концентрация муравьев протекает одинаково в обоих случаях. Следствием объединения семей-недоростков в одном гнезде оказывается последующий быстрый рост этого муравейника. Причин тут, видимо, немало: улучшение температурного режима в гнезде (в большом гнезде он успешнее выдерживается), повышение конкурентоспособности семьи и, что, наверное, самое главное, восстановление естественной структуры семьи. По-видимому, мы не ошибемся, если назовем подразделение большой семьи ряда видов на колонны обязательным условием их нормального существования и развития. Семья, состоящая из одной колонны, - всего лишь начальный этап жизни муравейников многих видов.
А что же происходит в тех случаях, когда "разбежавшиеся" колонны неопределенно долго остаются равными по численности? Этот вариант тоже ведь не исключен. Встречается и такое: два муравейника растут рядом, связанные обменной дорогой. Со временем в каждом из них возникают новые колонны, иногда дело доходит и до образования отводков. А давняя дорога продолжает существовать. Впрочем, не бесконечно долго. В конце концов муравейники-близнецы обособляются, и дорога между ними зарастает травой.
Таким образом, каждая колонна настолько индивидуальна и автономна, что всегда может начать самостоятельное существование. Но все-таки одноколонный муравейник, по-видимому, нерационален. Именно поэтому колонны образуются уже в небольших муравейниках. Общая численность населения такого гнезда находится на уровне 200 тыс., т. е. даже меньше численности одной колонны процветающего муравейника.
Имеется масса видов с небольшими семьями, которые никогда не достигают уровня, необходимого для формирования колонн. Сюда относятся муравьи с наиболее простой организацией и зачастую невыраженной структурой семьи. Для некоторых видов сама возможность образования внутрисемейных группировок особей открыта, но реализуется только в исключительных случаях. Есть и такие виды, у которых достижение семьей определенной (пороговой) численности служит сигналом к разделению ее пополам (гесмозису). Примерно так, как это происходит у медоносной пчелы. Только события, естественно, разворачиваются в гнезде или на поверхности почвы. Гесмозис у муравьев рода афеногастер описан французским энтомологом А. Леду в 1973 г. и рода мономориум - индийскими мирмекологами супругами Сеансами в 1971 г.
При делении старая самка вместе с частью рабочих покидает свой муравейник и основывает новый. В оставшемся гнезде из отложенных старой самкой яиц выкармливаются молодые самки, одна из которых после оплодотворения начинает яйцекладку. Эти виды монотонные, поэтому гесмозис связан с обеспечением самками обеих частей делящейся семьи. Этим обусловлено столь значительное сходство гесмозиса у данных видов муравьев и у пчел. Но гесмозис происходит и у некоторых полигинных муравьев, например у повсеместно распространенных у нас мирмика. Деление семьи мирмика, прослеженное А. В. Демченко, происходило по упрощенной схеме: самки, так же как и рабочие, делятся на две группы, после чего одна из группировок покидает старое гнездо и переходит в новое, сооружение которого начинается заблаговременно частью рабочих, муравьев.
Здесь, как и в гнездах формика, длительный процесс наращивания численности ведет к вызреванию в недрах процветающей семьи более мелких и соизмеримых с видовым оптимумом объединений индивидов. Различия в том, что семья не усложняется структурно, а распадается на две. Это происходит, очевидно, как следствие относительно слабых внутрисемейных связей. Отсутствует та могучая центростремительная сила, которая, как мы видели на примере рыжих лесных муравьев, не только доминирует над автономными силами колонн внутри гнезда, но и способна удержать колонны в рамках общей системы даже при его распадении. Вероятно, развитие механизмов, обеспечивающих сохранение целостности семьи при делении семьи на колонны, является тем эволюционным достижением, который по своему значению для дальнейшей судьбы муравьев намного превосходит даже сам принцип деления.
К этому вопросу мы еще вернемся, а пока лишь отметим, что в любом варианте деление общины происходит по дихотомическому принципу, что образующиеся при этом новые семьи или колонны - организационно завершенные и жизнеспособные единицы, способные к самостоятельному существованию. И, наконец, первая задача, которую решает в такой ситуации семья, - это оптимизация своей численности и организационной структуры. Образование новых муравейников в стратегию муравейника не входит, а является следствием решения первой задачи.
Необходимо отметить, что образование колонн резко повышает жизнеспособность каждого муравейника и комплексов муравейников. Это особенно важно для видов, сооружающих наземные купола из легких строительных материалов. Такие гнезда не защищают их обитателей от крупных млекопитающих: медведей, барсуков, кабанов, - а, наоборот, привлекают их. В этих условиях рассредоточение колонн по отдельным гнездам - единственное средство пассивной защиты, которое могут противопоставить муравьи. Небольшие муравейники звери почти не трогают. В результате одним из следствий массовых разорении муравейников кабанами оказывается увеличение общего числа гнезд при уменьшении их средних размеров.
В арсенале ряда видов имеются и некоторые другие средства выхода из критической ситуации, когда семья разрастается до таких размеров, что нормальное существование ее как единой системы индивидов становится невозможным. Речь об этом пойдет в последнем разделе данной главы.
Семья как система колонн
Итак, можно охарактеризовать семью муравьев как систему, состоящую из нескольких идентичных по составу и сравнимых по численности особей подсистем - колонн, которые имеют постоянный состав, занимают определенные части гнезда. Эти части (секции, секторы) подобны друг другу. Кормовой участок, используемый каждой колонной, также постоянен.
Колонны сравнимы по численности, и их отношения строятся не на иерархическом принципе, а на многостороннем равнодействии всех входящих в систему подсистем. Возможность проявления иерархических отношений исключается, так как вслед за непомерным усилением одной из колонн неотвратимо следует либо разделение этой колонны на две, либо выделение части ее населения в отводок. Муравейник действует как целостная система, хотя какие-либо специальные образования, координирующие рост и жизнедеятельность колонны или управляющие ими как подсистемами целого, отсутствуют.
Перечисленные свойства присущи особому координационному типу систем, представляющему собой довольно низкий уровень интеграции. На этом уровне в наименьшей степени проявляется зависимость подсистем друг от друга и от системы в целом, что мы и видим на примере муравьев. Общесемейные реакции формируются путем прямого сопоставления колонн между собой и каждой из них со всеми остальными.
Однако имеются и некоторые серьезные отличия системных свойств муравейника от традиционных координационных систем, например популяции. Из них стоит выделить три.
Во-первых, это ограниченное число подсистем, входящих в муравейник. Обычно 2-5, максимально 12, в то время как в любой благополучной популяции тысячи и миллионы индивидов. Немногочисленность подсистем повышает удельный вес каждой из них, делает значимым для состояния семьи любое изменение их количества. Одновременно функциональные связи между всеми колоннами становятся обязательными и регулярными. Прекращение таких связей означает выпадение колонны из семьи, чего в нормальных условиях не бывает. Все это приводит к значительно большей организованности муравейника как одного из вариантов координационных систем.
Во-вторых, колонны объединяются не только функциональными связями, но и общим местом обитания - гнездом, которое является основным выражением интеграции их деятельности. В общем гнезде регулярные связи всех колонн обретают важное дополнительное свойство: они осуществляются по строго определенным путям. И если в муравейнике нет координирующего функционального образования, какой-нибудь колонны-координатора, то там имеется определенная зона (внутренний конус или группа поверхностных ходов и камер), где сосредоточены основные пути координации. Это дополняет вытекающие из первого положения системные возможности семьи муравьев, еще более укрепляя ее организационную целостность, делая ее функционально более эффективной.
В-третьих, система контролирует изменение числа колонн уже на начальном этапе, регулируя их численность и размеры. Благодаря этому появилась возможность сопряженного устойчивого роста подсистем и семьи как объединяющего их целого. Почти стихийный рост колонн преобразуется в результате их взаимодействия в зарегулированный рост семьи. На определенном уровне численности появляется стабильность. Ее обеспечивает высокая степень организации муравьиной общины. Без четкой организации существование муравьев в таких количествах было бы просто невозможно.
Для более полной характеристики муравейника как системы требуется привлечение данных о системных свойствах самих колонн, т. е. об их поведенческой структуре. Этот чрезвычайно емкий и очень интересный вопрос не входит в нашу задачу. Однако стоит упомянуть, что поведенческая структура колонн достаточно сложна и, как правило, более видоспецифична. Все моменты, связанные с разнообразием каст и размерных групп муравьев, со сменой профессий и взаимодействием профессиональных групп между собой, добычей пищи и ритмикой общей активности, с особенностями поведения муравьев в гнезде и на участке, регулируются в рамках отдельной колонны.
Об организационной структуре муравейников известно еще очень мало. Многие вопросы требуют тщательной разработки, другие - едва лишь затронуты исследователями. Ограничен и круг видов, используемых в качестве модельных для этих целей. Но все-таки уже сейчас можно, хотя бы в первом приближении, представить себе, сколь сложна и совершенна организация семьи муравьев, насколько своеобразен их мир, преодолевший не одно, казалось бы, непреодолимое препятствие - мир, где собранные по крупинкам достижения индивидов синтезированы в строгие и действенные формы организации.
Поликалия у муравьев
Для рациональной организации семьи и при перенаселенности гнезда у муравьев наблюдается не только образование колонн. В арсенале ряда видов имеются некоторые другие возможности выхода из критической ситуации. В настоящее время мы имеем сведения еще о двух типах внутрисемейной организации - это поликалия* и диффузные гнезда*.
Поликалия - обитание одной семьи в нескольких взаимосвязанных гнездах - очень широко распространена у муравьев, в первую очередь среди видов, в семьях которых имеется только одна самка. Помимо основного, самого крупного гнезда муравьи сооружают вспомогательные, находящиеся на некотором расстоянии от основного и выполняющие различные функции. Эти гнезда гораздо меньше по размерам и могут существовать различное время: несколько дней, в течение одного сезона, ряда лет или всего срока существования семьи.