Получение рекомбинантных ДНК и их амплификация.
При получении рекомбинантных ДНК выделяют эти молекулы из двух разных источников. Каждую из них в отдельности фрагментируют, используя одну и ту же рестриктазу. После процедуры нагревания и медленного охлаждения смеси полученных фрагментов, наряду с исходными молекулами ДНК образуются и рекомбинантные, состоящие из участков ДНК, первоначально принадлежавших разным образцам. Используя технику рекомбинантных ДНК, удаётся исследовать варианты генов, ответственных за развитие многих заболеваний. Этим способом могут быть идентифицированы различные мутации.
Для получения значительных количеств рекомбинантного генетического материала проводят клонирование ДНК, предполагающее встраивание нужного фрагмента ДНК в векторную молекулу, Вектор обеспечивает проникновение этой рекомбинантной ДНК в бактериальные клетки. При размножении трансформированных бактерий происходит увеличение числа копий введенного фрагмента ДНК, а также синтез не свойственных бактериальной клетке, но весьма ценных для человека белковых продуктов. Таким способом получают вакцины, инсулин, гормон роста, факторы свертывания крови и др.
Работа с нуклеотидными последовательностями требует наличия достаточного количества материала для исследования. Поэтому фрагменты ДНК предварительно амплифицируют (увеличивают количество). Метод полимеразной цепной реакции (ПЦР), предложенный в 1983 г. Карри Муллисом, позволяет подвергать специфической амплификации в условиях in vitro любые образцы ДНК.
Полимеразная цепная реакция протекает в три стадии:
1. Денатурация. Инкубационную смесь, в которой содержится образец нужной ДНК, нагревают до температуры 90°С. При этом в течение 15 секунд происходит разрушение слабых водородных связей между нитями ДНК, и из одной двухцепочечной молекулы образуется две одноцепочечные.
2. Гибридизация праймеров. Температуру снижают до 50°С. При этом происходит гибридизация цепей ДНК с праймерами. Эта стадия обычно протекает 30 секунд.
3. Полимеризация. Инкубационную смесь нагревают до 70°С. При такой температуре полимераза удлиняет оба праймера с их 3'-концов. Праймеры дорастают до размеров матрицы. Этот процесс протекает в течение 90 секунд. В результате количество ДНК удваивается.
Процедуру проводят в автоматическом режиме в приборе – термоциклере (циклизаторе, амплификаторе). Это устройство позволяет задавать нужное количество циклов и выбирать оптимальные временные и температурные параметры. С помощью ПЦР можно получить достаточное количество копий участков ДНК, в которых предполагаются присутствие мутаций, полиморфизм сайтов, можно проводить ДНК-диагностику инфицированности пациентов вирусными, бактериальными и грибковыми возбудителями болезней.
Глава 8. Введение в метаболизм
Обмен веществ или метаболизм – это совокупность химических реакций в организме, которые обеспечивают его веществами и энергией, необходимыми для жизнедеятельности. Процесс метаболизма, сопровождающийся образованием более простых соединений из сложных, обозначают термином - катаболизм. Процесс, идущий в обратном направлении и приводящий, в конечном счете, к образованию сложного продукта из относительно более простых – анаболизм. Анаболические процессы сопровождаются потреблением энергии, катаболические – высвобождением.
Анаболизм и катаболизм не являются простым обращением реакций. Анаболические пути должны отличаться от путей катаболизма хотя бы одной из ферментативных реакций, чтобы регулироваться независимо, и за счет контроля активности этих ферментов регулируется суммарная скорость распада и синтеза веществ. Ферменты, которые определяют скорость всего процесса в целом, называются ключевыми.
Более того, путь по которому идет катаболизм той или иной молекулы, может быть непригодным для ее синтеза по энергетическим соображениям. Например, протекающие в печени расщепление глюкозы до пирувата представляет собой процесс, состоящий из 11 последовательных стадий, катализируемых специфическими ферментами. Казалось бы, синтез глюкозы из пирувата должен быть простым обращением всех этих ферментативных стадий её распада. Такой путь представляется на первый взгляд и самым естественным, и наиболее экономичным. Однако в действительности биосинтез глюкозы (глюконеогенез) в печени протекает иначе. Он включает лишь 8 из 11 ферментативных стадий, участвующих в ее распаде, а 3 недостающие стадии заменены в нем совсем другим набором ферментативных реакций, свойственным только этому биосинтетическому пути. Кроме того, реакции катаболизма и анаболизма часто разделены мембранами и протекают в разных компартментах клеток.
Таблица 8.1. Компартментализация некоторых метаболических путей в гепатоците
| Компартмент | Метаболические пути |
|---|---|
| Цитозоль | Гликолиз, многие реакции глюконеогенеза, активация аминокислот, синтез жирных кислот |
| Плазматическая мембрана | Энергозависимые транспортные системы |
| Ядро | Репликация ДНК, синтез различных видов РНК |
| Рибосомы | Синтез белка |
| Лизосомы | Изоляция гидролитических ферментов |
| Комплекс Гольджи | Образование плазматической мембраны и секреторных пузырьков |
| Микросомы | Локализация каталазы и оксидаз аминокислот |
| Эндоплазматическая сеть | Синтез липидов |
| Митохондрии | Цикл трикарбоновых кислот, цепь тканевого дыхания, окисление жирных кислот, окислительное фосфорилирование |
Метаболизм выполняет 4 функции:
1. снабжение организма химической энергией, полученной при расщеплении богатых энергией пищевых веществ;
2. превращение пищевых веществ в строительные блоки, которые используются в клетке для биосинтеза макромолекул;
3. сборка макромолекулярных (биополимеры) и надмолекулярных структур живого организма, пластическое и энергетическое поддержание его структуры;
4. синтез и разрушение тех биомолекул, которые необходимы для выполнения специфических функций клетки и организма.
Метаболический путь – это последовательность химических превращений конкретного вещества в организме. Промежуточные продукты, образующиеся в процессе превращения, называют метаболитами, а последнее соединение метаболического пути – конечным продукт. Примером метаболического пути является гликолиз, синтез холестерина.
Метаболический цикл – это такой метаболический путь, один из конечных продуктов которого идентичен одному из соединений вовлеченных в этот процесс. Наиболее важными в организме человека метаболическими циклами являются цикл трикарбоновых кислот (цикл Кребса) и орнитиновый цикл мочевинообразования.
Почти все метаболические реакции в конечном итоге связаны между собой, поскольку продукт одной ферментативной реакции служит субстратом для другой, которая в данном процессе играет роль следующей стадии. Таким образом, метаболизм можно представить в виде чрезвычайно сложной сети ферментативных реакций. Если поток питательных веществ в какой-нибудь одной части этой сети уменьшится или нарушится, то в ответ могут произойти изменения в другой части сети, для того чтобы это первое изменение было как-то уравновешено или скомпенсировано. Более того, и катаболические и анаболические реакции отрегулированы таким образом, чтобы они протекали наиболее экономично, то есть с наименьшей затратой энергии и веществ. Например, окисление питательных веществ в клетке совершается со скоростью, как раз достаточной для того, чтобы удовлетворить ее энергетические потребности в данный момент.
Специфические и общие пути катаболизма
В катаболизме различают три стадии:
1. Полимеры превращаются в мономеры (белки – в аминокислоты, углеводы в моносахариды, липиды – в глицерол и жирные кислоты). Химическая энергия при этом рассеивается в виде тепла.
2. Мономеры превращаются в общие продукты, в подавляющем большинстве в ацетил-КоА. Химическая энергия частично рассеивается в виде тепла, частично накапливается в виде восстановленных коферментных форм (НАДН, ФАДН2), частично запасается в макроэргических связях АТФ (субстратное фосфорилирование).
1-ая и 2-ая стадии катаболизма относятся к специфическим путям, которые уникальны для метаболизма белков, липидов и углеводов.
3. Заключительный этап катаболизма, сводится к окислению ацетил-КоА до СО2 и Н2О в реакциях цикла трикарбоновых кислот (цикла Кребса) – общий путь катаболизма. Окислительные реакции общего пути катаболизма сопряжены с цепью тканевого дыхания. При этом энергия (40–45%) запасается в виде АТФ (окислительное фосфорилирование).
В результате специфических и общих путей катаболизма биополимеры (белки, углеводы, липиды) распадаются до СО2, Н2О и NH3, которые являются основными конечными продуктами катаболизма.
Метаболиты в норме и при патологии
В живой клетке ежесекундно образуются сотни метаболитов. Однако их концентрации поддерживаются на определенном уровне, который является специфической биохимической константой или референтной величиной. При болезнях происходит изменение концентрации метаболитов, что является основой биохимической лабораторной диагностики. К нормальным метаболитам относят глюкозу, мочевину, холестерол, общий белок сыворотки крови и ряд других. Выход концентрации этих веществ за пределы физиологических норм (повышение либо снижение) говорит о нарушении их обмена в организме. Более того, ряд веществ в организме здорового человека обнаруживается только в определенных биологических жидкостях, что обуславливается спецификой их метаболизма. Например, белки сыворотки крови в норме не проходят через почечный фильтр и, соответственно, не обнаруживаются в моче. Но при воспалении почек (гломерулонефрите) белки (в первую очередь альбумины) проникают через капсулу клубочка, появляются в моче – протеинурия и трактуются как патологические компоненты мочи.
Патологическими метаболитами являются миеломные белки (белки Бенс-Джонса), парапротеины при макроглобулинемии Вальденштрема, накопление аномального гликогена при гликогенозах, разнообразных фракций сложных липидов при сфинголипидозах и т.д. Они обнаруживаются только при болезнях и для здорового организма не характерны.
Уровни изучения обмена веществ
Уровни изучения обмена веществ:
1. Целый организм.
2. Изолированные органы (перфузируемые).
3. Срезы тканей.
4. Культуры клеток.
5. Гомогенаты тканей.
6. Изолированные клеточные органеллы.
7. Молекулярный уровень (очищенные ферменты, рецепторы и т.д.).
Довольно часто для изучения метаболизма используют радиоактивные изотопы (H, P, C, S, O), которыми помечают вещества, вводимые в организм. Затем можно проследить клеточную локализацию этих веществ, определить период полураспада и их метаболические пути.
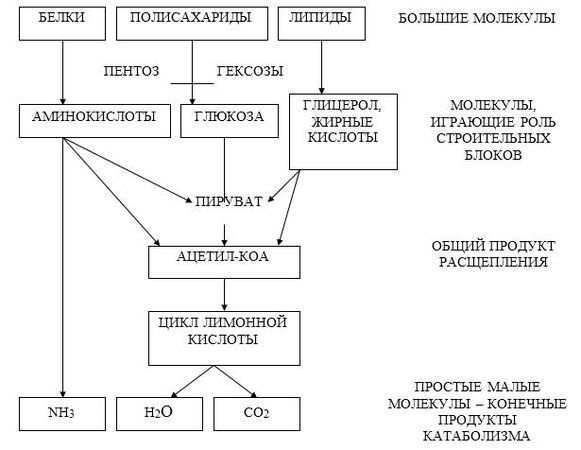
Рис. 8.1. Схема специфических и общих путей катаболизма
Глава 9. Биологические мембраны
Клетка представляет биологическую систему, основу которой составляют мембранные структуры, отделяющие клетку от внешней среды, формирующие ее отсеки (компартменты), а также обеспечивающие поступление и удаление метаболитов, восприятие и передачу сигналов и являющиеся структурными организаторами метаболических путей.
Согласованное функционирование мембранных систем – рецепторов, ферментов, транспортных механизмов помогает поддерживать гомеостаз клетки и в то же время быстро реагировать на изменения внешней среды.
Мембраны – нековалентные надмолекулярные структуры. Белки и липиды в них удерживаются вместе множеством нековалентных взаимодействий (кооперативных по характеру).
К основным функциям мембран можно отнести:
1. отделение клетки от окружающей среды и формирование внутриклеточных компартментов (отсеков);
2. контроль и регулирование транспорта огромного разнообразия веществ через мембраны (избирательная проницаемость);
3. участие в обеспечении межклеточных взаимодействий;
4. восприятие и передача сигнала внутрь клетки (рецепция);
5. локализация ферментов;
6. энерготрансформирующая функция.
Мембраны асимметричны в структурном и функциональном отношениях (углеводы локализуются всегда снаружи и их нет на внутренней стороне мембраны). Это динамичные структуры: входящие в их состав белки и липиды могут двигаться в плоскости мембраны (латеральная диффузия). Однако существует и переход белков и липидов с одной стороны мембраны на другую (поперечная диффузия, флип-флоп), которая происходит крайне медленно. Подвижность и текучесть мембран зависят от её состава: соотношениям насыщенных и ненасыщенных жирных кислот, а также холестерола. Текучесть мембраны тем ниже, чем выше насыщенность жирных кислот в фосфолипидах и чем больше содержание холестерола. Кроме того, для мембран характерна самосборка.
Общие свойства клеточных мембран:
1. легко проницаемы для воды и нейтральных липофильных соединений;
2. в меньшей степени проницаемы для полярных веществ (сахара, амиды);
3. плохо проницаемы для небольших ионов (Na, Cl и др.);
4. характерно высокое электрическое сопротивление;
5. асимметричность;
6. могут самопроизвольно восстанавливать целостность;
7. жидкостность.
Химический состав мембран.
Мембраны состоят из липидных и белковых молекул, относительное количество которых у разных мембран широко колеблется. Углеводы содержатся в форме гликопротеинов, гликолипидов и составляют 0,5%-10% веществ мембраны. Согласно жидкостно-мозаичной модели строения мембраны (Сенджер и Николсон, 1972г.) основу мембраны составляет двойной липидный слой, в формировании которого участвуют фосфолипиды и гликолипиды. Липидный бислой образован двумя рядами липидов, гидрофобные радикалы которых спрятаны внутрь, а гидрофильные группы обращены наружу и контактируют с водной средой. Белковые молекулы как бы растворены в липидном бислое и относительно свободно "плавают в липидном море в виде айсбергов на которых растут деревья гликокаликса".
Липиды мембран.
Мембранные липиды – амфифильные молекулы, т.е. в молекуле есть как гидрофильные группы (полярные головки), так и алифатические радикалы (гидрофобные хвосты), самопроизвольно формирующие бислой, в котором хвосты липидов обращены друг к другу. Толщина одного липидного слоя 2,5 нм, из которых 1 нм приходится на головку и 1,5 нм на хвост. В мембранах присутствуют три основных типа липидов: фосфолипиды, гликолипиды и холестерол. Среднее молярное отношение холестерол/фосфолипиды равно 0,3–0,4, но в плазматической мембране это соотношение гораздо выше (0,8–0,9). Наличие холестерола в мембранах уменьшает подвижность жирных кислот, снижает латеральную диффузию липидов и белков.
Фосфолипиды можно разделить на глицерофосфолипиды и сфингофосфолипиды. Наиболее распространенные глицерофосфолипиды мембран – фосфатидилхолины и фосфатидилэтаноламины. Каждый глицерофосфолипид, например фосфатидилхолин, представлен несколькими десятками фосфатидилхолинов, отличающихся друг от друга строением жирнокислотных остатков.
На долю глицерофосфолипидов приходится 2–8% всех фосфолипидов мембран. Наиболее распространенными являются фосфатидилинозитолы.
Специфические фосфолипиды внутренней мембраны митохондрий – кардиолипины (дифосфатидглицеролы), построенные на основе глицерола и двух остатков фосфатидной кислоты, составляют около 22% от всех фосфолипидов митохондриальных мембран.
В миелиновой оболочке нервных клеток в значительных количествах содержатся сфингомиелины.
Гликолипиды мембран представлены цереброзидами и ганглиозидами, в которых гидрофобная часть представлена церамидом. Гидрофильная группа – углеводный остаток – гликозидной связью присоединен к гидроксильной группе первого углеродного атома церамида. В значительных количествах гликолипиды находятся в мебранах клеток мозга, эпителия и эритроцитов. Ганглиозиды эритроцитов разных индивидуумов различаются строением олигосахаридных цепей и проявляют антигенные свойства.
Холестерол присутствует во всех мембранах животных клеток. Его молекула состоит из жесткого гидрофобного ядра и гибкой углеводородной цепи, единственная гидроксильная группа является полярной головкой.
Функции мембранных липидов.
Фосфо- и гликолипиды мембран, помимо участия в формировании липидного бислоя, выполняют ряд других функций. Липиды мембран формируют среду для функционирования мембранных белков, принимающих в ней нативную конформацию.
Некоторые мембранные липиды – предшественники вторичных посредников при передаче гормональных сигналов. Так фосфатидилинозитолдифосфат под действием фосфолипазы С гидролизируется до диацилглицерола и инозитолтрифосфата, являющихся вторичными посредниками гормонов.
Ряд липидов участвует в фиксации заякоренных белков. Примером заякоренного белка является ацетилхолинэстераза, которая фиксируется на постсинаптической мембране к фосфатитилинозитолу.
Белки мембран.
Мембранные белки отвечают за функциональную активность мембран и на их долю приходится от 30 до 70%. Белки мембран отличаются по своему положению в мембране. Они могут глубоко проникать в липидный бислой или даже пронизывать его – интегральные белки, разными способами прикрепляться к мембране – поверхностные белки, либо, ковалентно контактировать с ней – заякоренные белки. Поверхностные белки почти всегда гликозилированы. Олигосахаридные остатки защищают белок от протеолиза, участвуют в узнавании лигандов и адгезии.
Белки, локализованные в мембране, выполняют структурную и специфические функции:
1. транспортную;
2. ферментативную;
3. рецепторную;
4. антигенную.