3. нервный центр, в который входят один или несколько вставочных нейронов;
4. эфферентное нервное волокно (аксон эфферентного нейрона), по которому возбуждение направляется к органу.
В рефлекторной реакции всегда участвуют афферентные нейроны, передающие импульсы от рецепторов (например, проприорецепторов) исполнительного органа в ЦНС. С помощью обратной афферентации происходит коррекция ответной реакции нервными центрами, регулирующими данную функцию. Поэтому понятие "рефлекторная дуга" заменяется в настоящее время представлением о рефлекторном кольце, поскольку в функциональном отношении дуга замкнута и на периферии, и в центре беспрерывно циркулирующими во время работы органа нервными сигналами.
Простейшая рефлекторная дуга (моносинаптическая) состоит из двух нейронов: чувствительного и двигательного. Примером такого рефлекса является коленный рефлекс. Большинство рефлексов включают один или несколько последовательно связанных вставочных нейронов и называются полисинаптическими. Наиболее элементарной полисинаптической дугой является трехнейронная рефлекторная дуга, состоящая из чувствительного, вставочного и эфферентного нейронов. В осуществлении пищевых, дыхательных, сосудодвигательных рефлексов участвуют нейроны, расположенные на разных уровнях – в спинном, продолговатом, среднем и промежуточном мозге, в коре головного мозга.
Рефлексы возникают под влиянием специфических для них раздражителей, действующих на их рецептивное поле. Рецептивным полем рефлекса называется участок тела, содержащий рецепторы, раздражение которых всегда вызывает данную рефлекторную реакцию. Так, рефлекс сужения зрачка возникает при освещении сетчатки глаза, разгибание голени наступает при нанесении легкого удара по сухожилию ниже колена и т. д.
Нервные центры
Нервным центром называется функциональное объединение нейронов, обеспечивающее осуществление какого-либо рефлекса или регуляцию какой-либо определенной функции. Нейроны, входящие в нервный центр, обычно находятся в одном отделе ЦНС, но могут располагаться и в нескольких. Центр дыхания располагается в средней трети продолговатого мозга, центр мочеиспускания – в крестцовом, центр коленного рефлекса – в поясничном отделе спинного мозга. В осуществлении сложных рефлексов целостного организма принимают участие, как правило, не один, а многие центры, расположенные в разных отделах мозга, включая его высшие отделы. Например, в акте дыхания участвует не только центр дыхания в продолговатом мозге, но и нервные клетки варолиева моста, коры головного мозга и мотонейроны спинного мозга.
Особенности распространения возбуждения в ЦНС в основном определяются свойствами нервных центров:
1. Одностороннее проведение возбуждения. В ЦНС возбуждение может распространяться только в одном направлении: от рецепторного нейрона через вставочный к эфферентному нейрону, что обусловлено наличием синапсов.
2. Более медленное проведение возбуждения по сравнению с нервными волокнами. Промежуток времени от момента нанесения раздражения на рецептор до ответной реакции исполнительного органа называется временем рефлекса. Большая его часть тратится на проведение возбуждения в нервных центрах, где возбуждение проходит через синапсы. На выделение и диффузию медиатора в синапсе требуется промежуток времени в 1,5–2 мс (синоптическая задержка). Чем больше нейронов в рефлекторной дуге, тем продолжительнее время рефлекса.
3. Суммация возбуждений (или торможения). Нервные центры могут суммировать афферентные импульсы, что проявляется в усилении рефлекса при увеличении частоты раздражений или числа раздражаемых рецепторов. Различают два вида суммации: временная суммация – если импульсы приходят к нейрону по одному и тому же пути через один синапс с коротким интервалом, то происходит суммирование ВПСП на постсииаптической мембране и она деполяризуется до уровня, достаточного для генерации ПД; пространственная суммация связана с суммированием ВПСП, возникающих одновременно в разных синапсах одного нейрона. Оба вида суммации происходят в области аксонного холмика, где и генерируется ПД.
4. Конвергенция. В нервном центре несколько клеток могут передавать импульсы к одному нейрону, т. е. возбуждения конвергируют на нем. Конвергенция может быть результатом прихода возбуждающих или тормозных входных сигналов от различных источников. Так, моторные нейроны спинного мозга могут получать импульсы: от периферических нервных волокон, входящих в спинной мозг; волокон, соединяющих сегменты спинного мозга; кортикоспинальных волокон от коры мозга; тормозных путей от ретикулярной формации. В результате конвергенции происходит суммация импульсов от этих источников и возникает ответ, являющийся суммарным эффектом разнородной информации.
5. Дивергенция и иррадиация. Возбуждение даже единственного нервного волокна, по которому импульсы поступают в нервный центр, может послужить причиной возбуждения множества выходящих из центра нервных волокон. Морфологическим субстратом широкого распространения импульсов (иррадиации) возбуждения является ветвление аксонов и наличие большого числа вставочных нейронов в пределах центра.
6. Облегчение и окклюзия. На нейронах нервных центров оканчиваются не только волокна их собственных афферентных входов. Каждый из них получает веточки от афферентов соседнего центра, что может обусловливать развитие окклюзии (закупорки) или, наоборот, облегчения.
Феномен окклюзии состоит в том, что количество возбужденных нейронов при одновременном раздражении афферентных входов обоих нервных центров оказывается меньше, чем арифметическая сумма возбужденных нейронов при раздельном раздражении каждого афферентного входа в отдельности. Явление окклюзии приводит к снижению силы суммарной ответной реакции. Феномен центрального облегчения характеризуется противоположным эффектом. На облегчении основано проторение пути – распространение возбуждения не по той цепи нейронов, по которой информация не поступала ранее, а через нейроны, уже облегченные первым раздражением.
7. Трансформация ритма возбуждений. Если сопоставить частоту импульсов в задних (чувствительных) и передних (двигательных) корешках спинного мозга при раздражении рецепторов, то обычно их ритм не совпадает. Центры способны как снижать, так и повышать ритмы возбуждений, поступающих от рецепторов.
8. Рефлекторное последействие. Продолжительность рефлекса всегда больше, чем время раздражения, так как возбуждение в нервных центрах сохраняется в течение некоторого времени после прекращения действия раздражителя. Это объясняется тем, что вставочные нейроны в центре образуют замкнутые цепи ("нейронные ловушки"), по которым ПД могут длительно циркулировать.
9. Высокая чувствительность к недостатку кислорода. Уменьшение доставки к клеткам мозга кислорода быстро ведет к тяжелым расстройствам деятельности ЦНС и гибели нейронов. Кратковременное нарушение кровоснабжения мозга вследствие временного спазма его сосудов или падения давления крови приводит к потере сознания – обмороку. Своевременно принятые меры по восстановлению кровоснабжения мозга (нашатырный спирт, кофеин, горизонтальное положение тела и др.) выводят больного из обморока.
10. Высокая чувствительность к химическим веществам объясняется большим числом синапсов. На одном нейроне могут рас-полагаться синапсы, обладающие чувствительностью к различным химическим веществам. Подбирая фармакологические препараты, которые избирательно блокируют одни синапсы, оставляя другие в рабочем состоянии, можно корректировать реакции организма.
11. Низкая функциональная подвижность (лабильность) и высокая утомляемость. Нервные центры, как и синапсы, обладают низкой функциональной подвижностью и быстрой утомляемостью в отличие от нервных волокон, которые считаются практически неутомляемыми и имеют высокую лабильность.
12. Посттетаническая потенциация – явление усиления рефлекторного ответа после длительного ритмического раздражения нервного центра. Это связано с сохранением ВПСП на нейронах центра в течение некоторого времени, что облегчает проведение последующих возбуждений через синапсы.
13. Тонус нервных центров. Регистрация биоэлектрической активности ЦНС даже при отсутствии раздражений показывает, что многие нервные центры (составляющие их нейроны) постоянно генерируют импульсы. Эта импульсация поступает к рабочим органам и свидетельствует о существовании некоторого постоянного тонического возбуждения нервных центров.
14. Пластичность. Нервные центры обладают способностью изменять собственное функциональное назначение и расширять свои функциональные возможности, т. е. существенно модифицировать картину осуществляемых рефлекторных реакций. Пластичность нервных центров тесно связана с изменением эффективности или направленности связей между нейронами.
Торможение в центральной нервной системе и его виды
Нормальная деятельность отдельных нервных центров и ЦНС в целом может осуществляться лишь при обязательном участии тормозных процессов.
Торможение в ЦНС – это активный процесс, проявляющийся в подавлении или ослаблении возбуждения. Явление торможения в центральной нервной системе было открыто И. М. Сеченовым в 1862 г. в эксперименте на лягушке, у которой перерезали мозг на уровне зрительных бугров и удалили полушария головного мозга. После этого измеряли время рефлекса отдергивания задних лапок при погружении их в раствор серной кислоты. Этот рефлекс осуществляется спинномозговыми нейронами и его время служит показателем возбудимости нервных центров. Если на область зрительных бугров наложить кристалл хлорида натрия, то время рефлекса увеличивается, т. е. в области зрительных бугров имеются центры, оказывающие тормозящее влияние на спинномозговые рефлексы. Торможение может наступить не только в результате непосредственного воздействия на нервные центры, но и на рецепторы. Гольц показал, что рефлекс отдергивания одной лапки может быть заторможен более сильным раздражителем – сдавливанием другой лапки лягушки пинцетом. В данном случае торможение развивается в результате встречи двух возбуждений в ЦНС, т. е. если в ЦНС поступают импульсы из разных рецептивных полей, то более сильные раздражения угнетают слабые и рефлекс на последние тормозится. Таким образом, процесс торможения тесно связан с процессом возбуждения.
Классификация видов торможения
Торможение в ЦНС можно классифицировать по различным признакам (рис.9): электрическому состоянию мембраны (гипер-поляризационное и деполяризационное); отношению к синапсу (постсинаптическое и пресинаптическое); нейрональной организации (поступательное, возвратное, латеральное).
Торможение мембраны обычно является гиперполяризационным, тормозной медиатор увеличивает проницаемость мембраны для ионов калия или хлора (возникает ТПСП) и мембрану труднее довести до критического уровня деполяризации (при котором нейрон генерирует ПД). В нейронных сетях коры тормозная функция принадлежит в основном ГАМК-эргическим вставочным нейронам. Эти тормозные нейроны имеют относительно короткие проекции, поэтому их влияние ограничено локальными областями коры. Ингибирующее действие ГАМК осуществляется через ГАМКрецепторы А-типа, которые изменяют проницаемость мембраны для ионов хлора, что приводит к гиперполяризации мембраны. Есть данные, что ТПСП могут возникать и за счет взаимодействия ГАМК с рецепторами В-типа, которые связаны с калиевыми каналами. Деполяризация мембраны тоже может привести к торможению, если она становится чрезмерно длительной (застойной), такая деполяризация сопровождается инактивацией натриевых каналов, т.е мембрана теряет свою возбудимость и не в состоянии ответить на приход новых возбуждений генерацией ПД.
Постсинаптическое торможение
Это основной вид торможения, заключается в том, что в нервных окончаниях тормозящих нейронов под влиянием приходящего по аксону импульса выделяется медиатор, который гиперполяризует постсинаптическую мембрану другого возбуждающего нейрона. В мембране последнего деполяризация не может достигнуть критического уровня, и поэтому возбуждение по нейрону не распространяется.
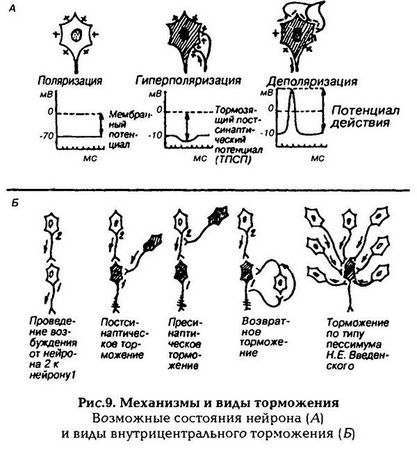
Пресинаптическое торможение
Локализуется в пресинаптических окончаниях, т. е. в разветвлениях (терминалях) возбуждающего нейрона. На этих терминалях располагаются окончания аксона тормозящего нейрона. При его возбуждении тормозной медиатор частично или полностью блокирует проведение возбуждения возбуждающего нейрона, и его влияние не передается на другой нейрон.
Поступательное торможение обусловлено тем, что на пути следования возбуждения встречается тормозной нейрон.
Возвратное торможение
Осуществляется вставочными тормозными клетками (клетками Реншоу). Аксоны мотонейронов часто дают коллатерали (ответвления), оканчивающиеся на клетках Реншоу. Аксоны клеток Реншоу оканчиваются на теле или дендритах этого мотонейрона, образуя тормозные синапсы. Возбуждение, возникающее в мотонейроне, распространяется по прямому пути к скелетной мускулатуре, а также по коллатералям к тормозящему нейрону, который посылает импульсы к мотонейронам и тормозит их. Чем сильнее возбуждение мотонейрона, тем сильнее возбуждаются клетки Реншоу и тем более интенсивно они оказывают свое тормозящее действие, что предохраняет нервные клетки от перевозбуждения.
Латеральное торможение
Является разновидностью возвратного торможения. Вставочные клетки могут формировать тормозные синапсы на соседних нейронах, блокируя боковые пути распространения возбуждения. В таких случаях возбуждение направляется по строго определенному пути. Этот вид торможения обеспечивает направленную иррадиацию возбуждения в ЦНС.
Пессимальное торможение
Для ЦНС свойственно пессимальное торможение, близкое по своему происхождению пессимуму Введенского в нервно-мышечных соединениях. Пессимальное торможение развивается в синапсах при разражениях высокой частоты и зависит от низкой лабильности синапсов. Данное явление связано с появлением при частых импульсах стойкой деполяризации постсинаптической мембраны, что нарушает распространение возбуждения и рефлекс тормозится. К пессимальному торможению особенно склонны вставочные нейроны спинного мозга и клетки ретикулярной формации. Пессимальное торможение предохраняет нервную клетку от перевозбуждения.
В нервной клетке может развиваться торможение после прекращения ее возбуждения – так называемое торможение вслед за возбуждением, возникающее в том случае, если после окончания возбуждения в клетке развивается сильная следовая гиперполяризация мембраны. Если в этот момент к клетке приходит новый импульс, то возникший посгсипаптический потенциал оказывается недостаточным для критической деполяризации мембраны.
Реципрокное (сопряженное) торможение.
Это явление, открытое Ч. Шеррингтоном, обеспечивает согласованную работу мышц-антагонистов, например, сгибателей и разгибателей конечностей, т. е. движение конечностей. При сгибании ног в коленном суставе развивается возбуждение в спинномозговом центре мышц-сгибателей и одновременно развивается торможение в нервном центре мышц-разгибателей. Наоборот, при разгибании, в нервном центре мышц-разгибателей наступает возбуждение, а в центре мышц-сгибателей – торможение. Такие взаимодействия названы реципрокным торможением. Реципрокное взаимодействие возникает и при более сложных двигательных актах, например при ходьбе. В этом случае происходит сгибание то одной, то другой ноги. Если в данный момент правое колено согнуто, то в центре сгибателей правой ноги развивается возбуждение, а в центре ее разгибателей – торможение. На левой стороне имеются противоположные взаимоотношения – центры разгибателей левой ноги возбуждены, а центры сгибателей заторможены. Во время следующего шага соотношения возбуждения и торможения в нейронах меняются в противоположном направлении. Чем сложнее двигательный акт, тем большее количество нейронов, регулирующих отдельные мышцы или их группы, находится в сопряженных соотношениях. Реципрокное торможение осуществляется при участии тормозящих вставочных нейронов спинного мозга. Реципрокные соотношения между отдельными центрами не постоянны и в типичной форме они выявляются лишь у спинальных животных. Реципрокное торможение может изменяться под влиянием нейронов, расположенных выше спинальных центров (особенно центров коры головного мозга). Например, обе ноги можно согнуть одновременно вопреки описанному выше сопряженному торможению центров сгибателей и разгибателей. Изменчивость взаимодействия между двигательными центрами обеспечивает сложнейшие движения человека во время разнообразной трудовой деятельности, сложных спортивных движений, танцев, игры на музыкальных инструментах и др.
Принципы координационной деятельности центральной нервной системы
Для осуществления сложных реакций необходима интеграция работы отдельных нервных центров. Большинство рефлексов являются сложными, последовательно и одновременно совершающимися реакциями. Рефлексы при нормальном состоянии организма строго упорядочены, так как имеются общие механизмы их координации. Возбуждения, возникающие в ЦНС, иррадиируют по ее центрам. Координация обеспечивается избирательным возбуждением одних центров и торможением других. Координация – это объединение рефлекторной деятельности ЦНС в единое целое, что обеспечивает реализацию всех функций организма.
Выделяют следующие основные принципы координации:
1. Принцип иррадиации возбуждений. Нейроны разных центров связаны между собой вставочными нейронами, поэтому импульсы, поступающие при сильном и длительном раздражении рецепторов, могут вызвать возбуждение не только нейронов центра данного рефлекса, но и других нейронов. Например, если раздражать у спинальной лягушки одну из задних лапок, слабо сдавливая ее пинцетом, то она сокращается (оборонительный рефлекс), если раздражение усилить, то происходит сокращение обеих задних лапок и даже передних. Иррадиация возбуждения обеспечивает при сильных и биологически значимых раздражениях включение в ответную реакцию большего количества мотонейронов.