В Атлантическом и Тихом океанах имеется по шесть крупномасштабных круговоротов, а в Индийском, северная часть которого подвержена существенным сезонным изменениям в связи со сменой муссонов,- четыре круговорота зимой северного полушария и три - летом. При этом в циклонических тропических и субполярных круговоротах (в них вода перемещается против часовой стрелки в северном полушарии и по стрелке - в южном) образуются зоны расхождения (дивергенции) вод, в которых развиваются восходящие токи воды, а в антициклонических субтропических круговоротах (направление вращения в них обратное по сравнению с циклоническими) поверхностные воды сходятся в центральных областях и погружаются там на глубину. Эти круговороты образуют гомологические пары, симметрично расположенные по отношению к экватору. Правда, эта симметрия (как и у большинства природных объектов) не обладает совершенством и полнотой, т. е. не идеальна и не касается всех признаков. Границы между круговоротами в открытом океане идут примерно в широтном направлении. Близ берегов зональное течение, общее для двух соседних круговоротов, раздваивается и между обеими крупномасштабными циркуляциями и берегом остаются треугольники не захватываемой ими воды - так называемые нейтральные области. В них существуют локальные круговороты меньшего масштаба.
В пределах каждого крупномасштабного круговорота набор водных масс по вертикали специфичен, а это означает, что в каждом круговороте имеется своя структура вод. Принято различать поверхностную, подповерхностную, промежуточную, глубинную и придонную водные массы, которые разделены слоями воды с повышенными градиентами свойств (в пределах каждой массы свойства не меняются или меняются очень постепенно). Выделяют два главных типа гидрологических структур - тропические, расположенные в тропиках и субтропиках, и субполярные, расположенные в умеренных широтах.
Для тропических структур характерны следующие водные массы и слои раздела между ними. Сверху находится поверхностный однородный (изотермический) слой, который характеризуется высокой температурой (20- 30°), мало меняющейся по глубине. Толщина этого слоя в среднем равна 50 - 100 м (в центральных частях субтропических круговоротов до 200 - 250 м). Под изотермическим слоем температура воды сравнительно резко падает, и ее градиент до глубины 400 - 500 м имеет порядок 0,1 град/м. Одновременно резко возрастает плотность воды. Под слоем повышенного градиента температура продолжает понижаться, хотя и более медленно. До глубины около 1000 - 1500 м (в зависимости от структуры) это понижение происходит, однако, быстрее, чем на еще больших глубинах. Этот слой называют "главным термоклином", и его нижняя граница выражена не очень четко. Глубже него вода отличается низкой температурой (около 2°), мало меняющейся с глубиной. Это так называемые "глубинные однородные воды".
Резкое расслоение толщи воды по температуре, а следовательно, и по плотности чрезвычайно характерно для тропиков. Плотность воды зависит, однако, и от солености, а в большей части тропической зоны выражен промежуточный слой повышенной солености на глубине нескольких сот метров, т. е. в верхней части главного термоклина. В результате главный термоклин распадается на два слоя: слой максимума солености до глубин 200 - 400 м и слой между ним и границей глубинных однородных вод, обозначаемый обычно как промежуточный минимум солености.
В субполярных водах весь столб воды сравнительно однороден по температуре. Главного термоклина там практически нет, и набор водных масс существенно отличается от характерного для тропической зоны. Поверхностный изотермический слой может нагреваться летом выше 10° (при толщине около 50 м) и охлаждаться зимой до 0 - 5° (иногда и до отрицательных значений температуры). Под этим слоем летом может существовать промежуточный холодный слой, в котором температура удерживается на уровне ее минимального зимнего значения; он может достигать глубины около 300 м. Промежуточный теплый слой с температурой на 1 - 2° выше, чем в предыдущем, располагается примерно до глубины 2000 м. Глубинные однородные воды мало отличаются по своим характеристикам от глубинных вод в тропических структурах.
Рассматривая вертикальную термическую структуру Мирового океана, нередко придают особое значение противопоставлению теплого поверхностного и холодного глубинного объемов водной толщи, называемых соответственно "термосферой" и "психросферой". Автор этих терминов - датский океанолог и ихтиолог А. Бруун принимал за границу таких слоев изотермическую поверхность 10°. Таким образом, в высоких широтах термосфера, естественно, отсутствует, в умеренных широтах она имеет сезонный характер, а в тропической зоне постоянна, но варьирует по толщине от 100 м в восточных районах океанов, характеризующихся подъемом глубинных вод, до 700 м в западных.
Каждый участок дна омывается водами того или иного слоя водной толщи, и поэтому дно по вертикали может подразделяться на те же участки, что и пелагиаль (тонкий слой воды у самого дна изучен недостаточно, и о его характеристиках судят в основном по характеристикам основного объема соответствующей водной массы). На дне океана выделяются, кроме того, участки (зоны), различающиеся режимом и скоростью осадконакопления, размерами частиц грунта и содержанием в нем органических веществ.
Почти все органическое вещество в океане первично продуцируется в приповерхностной зоне - в освещенных (эвфотических) слоях, где возможен фотосинтез, а затем разными путями проникает вглубь и служит той пищей, которая обеспечивает существование населения в лишенных света слоях воды и на дне. Роль органического вещества, создаваемого в лишенных света глубинах океана за счет первичных хемосинтетических процессов, невелика: оно составляет лишь доли процента от синтезируемого растениями.
Продукция фитопланктона определяется целым рядом факторов - интенсивностью солнечной радиации, содержанием биогенных солей (фосфатов, нитратов, силикатов), толщиной верхнего однородного слоя, прозрачностью воды и т. д. Главным лимитирующим показателем практически везде служит все же концентрация в эвфотической зоне биогенных солей, основной запас которых находится в более глубоких слоях воды. Распределение этих солей в открытом океане обусловлено в первую очередь динамическими факторами, вызывающими подъем глубинных вод в верхний слой. В соответствии с этим районы наибольшей первичной продукции расположены в умеренных широтах, где обогащение поверхностных слоев питательными веществами происходит в результате зимнего конвективного перемешивания, а также в некоторых районах тропической области, характеризующихся наиболее интенсивным подъемом "свежих" глубинных вод. К ним относятся районы поднятия вод (апвеллингов) в зонах действия сгонных ветров у берегов, дивергенции течений (особенно субэкваториальная дивергенция) и центральные участки некоторых локальных круговоротов. Наименьшими показателями продукции фитопланктона отличаются обширные по площади области опускания вод в антициклонических субтропических круговоротах. Ежегодная суммарная величина первичной продукции составляет, по современным представлениям, не менее 60 млрд т углерода. Продукция фитопланктона, представляющего собой самый низший трофический уровень в пищевых взаимосвязях населения пелагиали, служит основой общей биологической продуктивности океана и в конечном счете его промысловой продуктивности. Дальнейшее перераспределение органического вещества, синтезированного водорослями-продуцентами, осуществляется животными-консументами - растительноядным и хищным зоопланктоном, а затем нектоном - и проходит, таким образом, через несколько трофических уровней. При переходе на каждый последующий уровень расходуется большая часть энергии (такие потери часто достигают 90%), и это ограничивает число уровней: в холодных водах их 3 - 5, в тропиках - до 7. В одних пелагических сообществах, (их называют сбалансированными по трофическим циклам) продукция, ежегодно создаваемая на каждом уровне, нацело выедается хищниками следующего уровня, в других (несбалансированных) эта продукция длительно и существенно превышает выедание и ее избыток выносится течениями в другие участки или попадает на дно. Наименьшей сбалансированностью отличаются неритические сообщества, а максимальное приближение к сбалансированности имеет место в тропической части открытого океана, где наблюдается высокая стабильность в количественном соотношении организмов, стоящих на разных трофических уровнях.
Количественное распределение и продукция зоопланктона в океане в общих чертах соответствуют обилию растительной жизни. Наибольшая биомасса среднеразмерного зоопланктона ("сетного" зоопланктона, или мезопланктона) наблюдается летом в умеренных и высоких широтах (прежде всего во фронтальных зонах), а также в экваториальных районах и районах глобальных подъемов вод в восточных частях океанов, наименьшая биомасса - в зонах антициклонических субтропических круговоротов, так называемых "океанических пустынях". Наибольшая продукция мезопланктона также создается в субполярных водах обоих полушарий и в водах прибрежных и экваториальных апвеллингов, а иногда и в районах локальных продуктивных пятен.
Распределение биомассы океанического макропланктона и нектона тоже подчинено широтной и циркумконтинентальной зональности. Прослеживаются три полосы повышенной биомассы - в северных умеренных, экваториальных и южных умеренных широтах, причем в экваториальной зоне количественные показатели обилия снижаются в направлении с востока на запад в Тихом и Атлантическом океанах и с северо-запада на восток - в Индийском океане. Эти полосы перемежаются четырьмя полосами пониженной биомассы - арктической, двумя центральными (субтропическими) и антарктической, в которой биомасса все же значительно выше, чем в трех других. Во всех океанах значительная биомасса консументов высших трофических уровней (главным образом рыб) приурочена к нерито-океанической зоне, переходной между прибрежными и океанскими районами. В этих участках максимальная биомасса нектона наблюдается на западных окраинах океанов в зоне полярных фронтов и на их восточной периферии мористее районов пассатных апвеллингов и вдоль прилегающих фронтальных разделов. В Индийском океане, где северного полярного фронта и северо-восточного апвеллинга нет, повышенная биомасса наблюдается мористее районов Аравийского и Сомалийского сезонных апвеллингов. В экваториальных широтах количество макропланктона и нектона особенно велико на востоке океанов (продуктивные зоны Гвинейского и Панамского заливов и прилегающих вод) и в районах локальных апвеллингов Карибского моря - Мексиканского залива и австрало-азиатских морей.
Все перечисленные выше (а также некоторые другие) особенности открытого океана оказывают непосредственное влияние на пространственное распределение обитающих в его водах организмов, в том числе и рыб. Участки, различающиеся по своему населению, могут иметь границы, совпадающие с изобатами, изотермами, изогалинами, линиями равного содержания растворенного кислорода, первичной продукции, биомассы мезопланктона и т. д., однако чаще всего эти границы соответствуют разделам дискретных водных масс, на которые подразделяется водная толща. Поэтому многие исследователи считают, что деление океана на крупномасштабные биотопы - участки, имеющие однородное население, - в первую очередь определяется именно расчлененностью его объема (так называемая "гипотеза водных масс").
Границы водных масс в придонных горизонтах неплохо совпадают с основными формами рельефа дна. В вертикальном плане океан подразделяется поэтому на следующие зоны: верхнюю (от поверхности до глубины 100- 200 м), среднюю (до глубины 1000 - 1500 м), нижнюю (до глубины 3000 м), глубинную (до глубины 6000 м) и сверхглубинную (рис. 1). В классификации участков океанической среды обитания наибольшее значение имеет их разделение на пелагические биотопы, с одной стороны, и донно-придонные - с другой. В пелагиали верхняя зона соответствует поверхностным и подповерхностным водам, средняя - промежуточным, нижняя - глубинным, глубинная - придонным, а сверхглубинная - водам глубоководных желобов; на дне (в бентали) верхняя зона более или менее совпадает с (шельфом; средняя и нижняя - с материковым склоном, глубинная - с ложем океана, а сверхглубинная - с дном желобов. При этом нужно иметь в виду, что приведенные примерные глубины не могут рассматриваться как абсолютные показатели границ вертикальных зон: их положение сильно варьирует в разных районах Мирового океана.
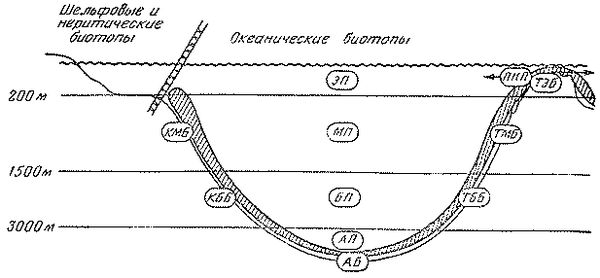
Рис. 1 . Вертикальная зональность океанических биотопов
Зоны водной толщи: эпипелагиаль (ЭП), мезопелагиаль (МП), батипелагиаль (БП), абиссопелагиаль (АП), псевдонеритнческий участок (ПНП); зоны донно-придонных биотопов (в субконтинентальной - С и талассной - Т модификациях, бентопелагические сообщества обозначены штриховкой): эпибенталь (ТЗВ), мезобенталь (КМБ и ТМБ), батибенталь (КББ и ТББ) и абиссобенталь (АБ)
Названные вертикальные зоны в пелагиали вполне традиционно именуются эпи-, мезо-, бати-, абиссо- и хадопелагиалью. Для бентали этих же зон в СССР обычно применяют названия сублитораль (на шельфе), "переходный горизонт" и батиаль (соответственно на верхней и нижней частях склона), абиссаль (на ложе океана) и ультраабиссаль (на дне глубоководных желобов). Им эквивалентны вертикальные зоны, названные Т. С. Рассом эпи-, мезо-, бати-, абиссо- и хадобенталью, и эта унифицированная терминология представляется более удобной для сопоставления биотопов дна и толщи воды (термин "батиаль" сохраняется при этом для обозначения всего биотопа материкового склона, т. е. идентичен понятию "мезобатибенталь").
Население пелагиали контактирует с населением бентали во всех вертикальных зонах в придонных слоях водной толщи, которые могут рассматриваться как биотоп вторичного сообщества - бентопелагического экотона (по определению К. В. Беклемишева, "экотон есть биотоп сообщества, состоящего из смеси видов двух соприкасающихся биоценозов"). В составе этого экотона в целом преобладают пелагические виды, а настоящие бентосные, как правило, находятся лишь на ранних стадиях жизненного цикла. В бентопелагиали имеются и специфические виды, и именно они составляют наиболее характерную часть ее населения. Это сообщество особенно четко обособлено в самой верхней вертикальной зоне (хорошо известное население неритической эпипелагиали), но оно существует во всем диапазоне океанских глубин. Бентопелагиаль, как и пелагиаль, является трехмерным биотопом и отличается в этом отношении от "плоской" двухмерной бентали, но объем придонной воды составляет ничтожную часть общего объема водной толщи, и в планетарном масштабе донный и придонный биотопы представляются тесно сопряженными (действительно, они не могут существовать в разделенном виде).
В дополнение к приведенной выше схеме вертикальной зональности А. П. Андрияшев выделил переуглубленные участки дна на шельфе - псевдобатиаль и изолированные от океанских глубин глубокие (более 3000 м) бассейны - псевдоабиссаль. Примеры псевдобатиального биотопа дают многочисленные депрессии на шельфе Антарктиды и в бассейне Северного Ледовитого океана, отделенные мелководьями от материкового склона; примерами псевдоабиссали (псевдоабиссобентали) являются Японское и Средиземное моря, соединяющиеся с океаном неглубокими проливами, обособленные котловины Полярного бассейна и т. д. Бесшельфовые подводные поднятия с глубинами порядка батиальных были выделены в талассобатиаль, и этот термин (также предложенный А. П. Андрияшевым) широко применяется в отечественной литературе. По аналогии с ним вершины подводных гор (часто их неправильно называют банками) с глубиной порядка 200 м и менее и "шельфы" малых островов были названы мной талассосублиторалью, или талассоэпибенталью.
Кроме того, над высокими подводными поднятиями и около них (а иногда, по-видимому, и в других участках со слабо сбалансированными трофическими циклами) могут возникать условия, в некоторой степени имитирующие особенности неритической зоны, и это позволяет обозначать такие участки как псевдонеритические (или талассоэпипелагические). Таким образом, в бентали а бентопелагиали Мирового океана три зоны - верхняя, средняя и нижняя - существуют в двух модификациях - субконтинентальной и талассной, а зона верхней пелагиали даже в трех - субконтииенталъной (неритической), собственно океанической и талассной (псевдонеритической). Из семи субконтинентальных биотопов к океаническим относятся, однако, только донные и придонные биотопы материкового склона (соответствующие модификации трех биотопов верхней зоны являются шельфо-неритическими) (табл. 1).
Таблица 1
Основные биотопы открытого океана